
The Primer Pheromones and Managing the Labor Pool – Part 2

March 30, 2011
The Primer Pheromones and Managing the Labor Pool
Part 2
Randy Oliver
ScientificBeekeeping.com
First published in American Bee Journal May 2010
In the first part of this article, I explained how the allotment of the hive labor pool was largely controlled through communication via the process of sharing protein-rich jelly via trophallaxis, plus non-feeding interchange of low volatility “primer” pheromones. I now continue with a summary of our current state of knowledge of the complex process of labor allotment within the hive.
The “Purpose” of the Hive Economy
Perhaps we should first define the “purpose” that drives the honey bee economy. The function of bees in nature is that they pollinate plants, but that’s just a convenient symbiotic collaboration that allows them to follow their true purpose (note that the fickle bee doesn’t care about any particular plant’s success—for example, they will abandon species with low-sugar nectar (such as pears) the moment that a more attractive nectar becomes available (as from apples)).
The purpose of the bee economy is that of successfully passing the workers’ genes into the next generations via drones or swarms—i.e., colony reproduction. So let’s look at how this works–starting with a fresh swarm. Its first order of business after moving into a new cavity is to build comb and store enough honey so that it can both make it through the winter and build up early enough the next spring to swarm before the main honeyflow. They will also generally need to replace their aging queen mother via supersedure. Come fall, they must then shift from foraging and broodrearing to the production of long-lived “winter bees.” Then in spring, they strive to madly expand their population, produce new queens and drones, and to send off one or more fresh swarms (colony reproduction). Not very romantic, but if you understand the bees’ purpose, you can better work with them, rather than against them.
Practical Tip: If you constantly keep in mind that the bees’ goals are invariably geared toward reproduction and winter survival, then you can use the bees’ innate behaviors to your advantage. Here’s a quote that I read to all my newbee beginners:
“The bees have their definite plan for life, perfected through countless ages, and nothing you can do will ever turn them from it. You can delay their work, or you can even thwart it altogether, but no one has ever succeeded in changing a single principle in bee-life. And so the best bee-master is always the one who most exactly obeys the orders from the hive”—from The Bee-master of Warrilow, Ticknor Edwards, 1920.
So here’s what’s interesting to me– the colony must strike a delicate balance between maintaining enough nurses to care for the brood and to produce jelly for the queen and foragers; plus delegate the optimal number of workers to the foraging force; yet maintain the right amount of mid-aged honey processors to support them. How the heck does the colony “know” how to allocate the right proportion of workers to each critical task? Not only that, but any established balance must be quickly readjusted when a honeyflow starts or ends!
So what are the proximate causes that regulate colony balance and response to the environment, not to mention the initiation of swarming or supersedure? At this point I’d like to closely investigate the interactions between pollen and nectar income, the transfer of jelly, and communication via pheromones.
A Model
On my first trip to Australia, a lady beekeeper cracked me up when she thanked me for “translating what the ‘boffins’ (researchers) say, into English.” I realize that by this point, this article is starting to sound a lot like boffin talk. So I’m going to take a bold step, and create a diagram that summarizes our current state of knowledge on the influence of the primer pheromones within the colony. In the model, I hope to illustrate how the “adaptable workforce” that I described in my last article is continually adjusted in response to colony needs. Please realize that this graphic is a very simplified version. It will be helpful for you to refer to it throughout the rest of this article.
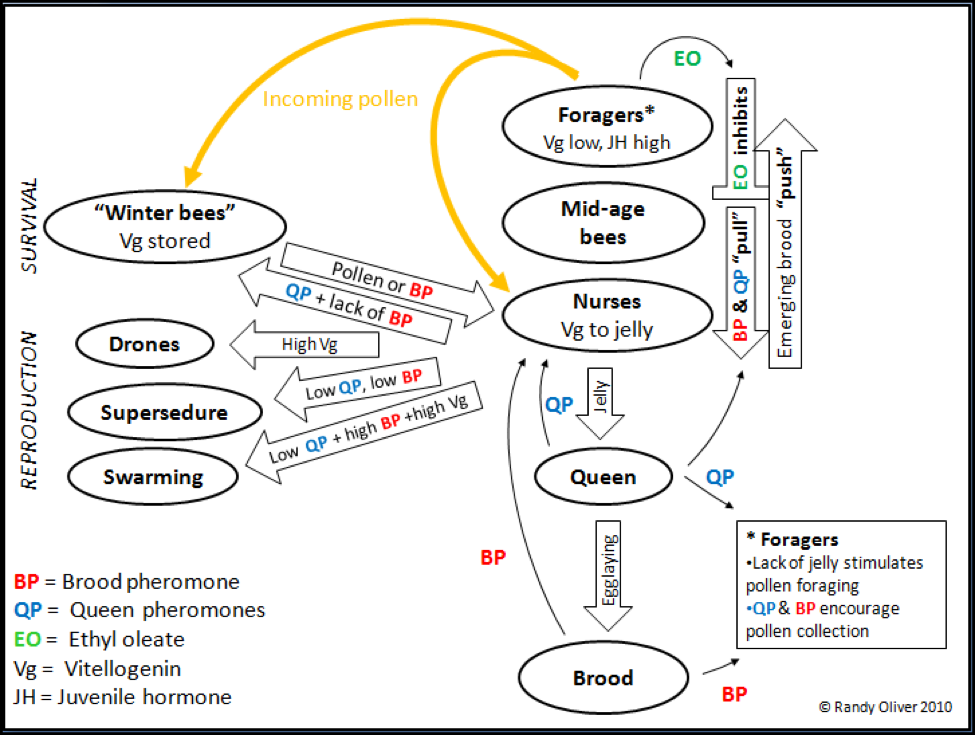
Figure 2. A simplified diagram of the transfer and effects of primer pheromones upon bee physiology, “aging,” and task behavior; and colony reproduction and wintering. Note the overriding importance of fresh pollen income, and the multiple effects of brood pheromone and queen pheromone. Also note the “push/pull” on mid-aged bees to maintain a balance between foragers and food processors.
Update March 2017: Keen-eyed beekeeper Blake Bennett pointed out to me that my article “Understanding Colony Buildup 7b” contradicts the influence of brood pheromone upon swarming. What I now realize is that I must separate “young larval pheromone” (e-beta-ocimene) from “older larva pheromone” and pupal pheromones, since it may be the abundance of pupal pheromone coupled with an absence of e-beta-ocimene that triggers the initiation of the swarm impulse.
OK, there’s a lot of information in the above graphic! But I feel that it is worth taking the time to study it carefully, in order to get a better feel of what’s going on in a hive. Let’s start with the most important input to the economy of the hive—a pollen flow.
The colony responds quickly to incoming pollen. A pollen flow is the signal to rouse diutinus bees (during winter or drought) out of their “survival” mode. They gorge on the precious protein, and start feeding jelly to the queen, who responds in turn by initiating egglaying. The queen continually announces her presence, behavior, location, and likely her state of fertility and hunger, by secreting queen mandibular pheromone, along with several lesser pheromones. I’m intentionally calling them collectively “queen pheromone” (QP).
As soon as the eggs hatch, hungry young larvae start pumping out “feed me” brood pheromone. This BP in turn causes the diutinus bees to shift from vitellogenin storage in their fat bodies, to mobilizing it into their blood, the better to produce jelly.
Practical application: races of bees from extreme climates (such as the Primorsky Russians or Africans) are far more responsive to changes in pollen flows than are those from Mediterranean climates (such as Italians). The keepers of Russian bees also tell me that Russians are less responsive to artificial pollen supplements than are Italians.
Since there is still not enough BP in circulation at this point to demand that very many bees take on nursing duties, the remaining “winter bees” are free to respond to the dances of pollen- and nectar-collectors, and are recruited into foraging. At the same time, the rising level of BP from the increasing amount of brood stimulates those foragers to collect more pollen (Pankiw 2004)!
As long as there is abundant brood (secreting BP) the nurses knows that there is a laying queen (due to BP and QP), and that they need to continue to devour pollen in order to make enough jelly to feed everyone. High titers of BP and QP “pull” against the maturation process of nurses into mid-age bees, keeping them in the broodnest (Toth 2005). (I want to give credit to Harris (2008a) and Johnson (2010) for the “push/pull” analogy).
Then as fresh adult bees emerge from their tighly-packed cells, they physically take up more than three times as much comb space as they did as pupae!
Practical application—predicting colony strength prior to almond pollination: Say that you’re inspecting your hives two weeks prior to bloom. For colonies wintered in cold/cool areas, this inspection occurs at the critical time when they are transitioning from their winter lull to serious spring buildup. At this time, the aging “winter bee” population is dying off at the rate of a half to full frame of bees during this two-week period (Harris, unpublished data), assuming that the colony is well fed, and not having problems with mites, nosema, viruses, etc.
Since you’ve only got two weeks, only existing sealed brood can possibly add to the adult population. So for the colony to grow in strength over the next two weeks, it would need to have sealed brood in excess of maybe a third to half a frame. Each full frame of brood (or equivalent) over that amount will increase colony strength by about three frames of bees. So, for example, a healthy 5-frame colony would need to have about 1½ full frames of sealed brood to make 8-frame grade two weeks later. (You can do the math for other colony strengths).
The resultant crowding as a consequence of worker emergence physically displaces the older nurses toward the cooler periphery of the broodnest. The newly-emerged bees engage in cell cleaning and capping, while they gorge on pollen in order to develop their brood food glands, and to mature their wing muscles in the warmth of the broodnest. In a few days, they begin to produce jelly and act as nurses.
Meanwhile, the displaced nurses are no longer exposed to as much BP or QP. As a consequence, these “mid-aged bees” now ignore the brood (not my job anymore!), but rather start paying attention to the other hive tasks at hand–packing pollen, receiving and processing nectar, building comb, cleaning debris, ventilating, and guarding.
***
Update 12/15/2016: A recent study is of interest: Eyer, M, et al (2017) Social regulation of ageing by young workers in the honey bee, Apis mellifera. Experimental Gerontology 87: 84–91. Their conclusion (paraphrased): The presence of young workers in broodless colonies can reduce the lifespan of older workers similar to the previously described effect of brood. However, the differential expression of all but one tested genes, suggests that young workers and brood influence ageing of adult nestmate workers via different physiological pathways.
Two things then occur to me:
- That the presence of young workers, but no brood, would signal that the colony is entering a population decline phase (due to lack of incoming pollen), and that the recently emerged workers would be transitioning to diutinus state. In that case, it would be adaptive for the colony to dump its older, food-consuming foragers, since there would be no more pollen to collect.
- Those recently-emerged workers, sensing no brood, would use all available protein to load their fat bodies (for sustained survival), and thus severely restrict the amount of jelly that they would feed to begging foragers. This lack of jelly feeding would accelerate the aging of those foragers.
The two phenomena above would be adaptive for a colony going into a pollen stress period, whether in summer or winter.
***
Mid-aged bees are physiologically fully developed, and in short order can engage in most any task. But most important, they serve as nectar receivers and processors to take the incoming nectar from the foragers. Any incoming nectar is transferred from foragers to receivers in the dance area near the entrance, who then carry it up to the top of the brood nest for immediate use by the nurses, or for processing any excess into stored honey. Should they not find empty cells in which to place nectar, they are forced to hold it in their crops—this storage stimulates their wax glands to go into production, which then allows them to build new comb.
Practical application: this is when you will see “white wax” or “whitening of the combs,” meaning that the hive needs more storage space (or that it is ready to draw foundation).
If there’s no immediate task at hand, mid-age bees just wait in reserve, or patrol around looking for something useful to do. Johnson (2009) proposes that such patrolling may serve two important functions: that of assessing colony needs, and maintaining a flux of eager workers ready anywhere in the hive to jump on any job that needs to be done.
This is an important point! As long as workers are nurses or mid-age bees, they can live for a long time. However, as soon as they start foraging, the clock starts ticking. Active foragers during an intense honey flow only live for two weeks at best. So it would be counterproductive for mid-age bees to transition to foragers unless they are truly needed. Better to wait and bide their time as long-lived “general labor.”
The bees have developed a clever way for the mid-age bees to tell if there are enough foragers—the foragers secrete a pheromone (ethyl oleate—EO; Leoncini 2004) that inhibits the mid-age bees’ maturation process (by keeping Vg high)! Whenever there are not enough foragers (honey flow on, pesticide kill, adult bee disease), the EO inhibition is relieved, and mid-age bees rush to fill the void.
In an expanding colony during springtime, there will be a low nurse/brood ratio, thus BP titers will remain high, so the colony population will be mostly “house bees.” However, once colony growth starts to peak, there is then a shift toward a higher nurse/brood ratio, BP titers drop, and there will be a large population of middle aged bees at the ready to take advantage of the main honeyflow.
Practical application: in order to produce the most honey, the beekeeper tries to time colony growth so that the population peaks just as the main honeyflow begins. This may be tricky, since the weather often does not cooperate. Successful management may involve appropriate stimulative feeding to promote growth, or splitting and swarm control to reign it in. In beekeeping, timing is critical!
Since foraging is the riskiest behavior for a bee (other than colony defense), workers simply stay busy in the safe confines of the hive until they are needed to forage. But what happens when they are stressed, and somehow sense that they may not live much longer? In that case, they get their sick butts out of the hive, and work themselves to death, even in conditions that healthy bees wouldn’t normally fly in! (Amdam 2004, Lin 2004, Higes 2008, Lourenco 2009). In a review by Tofilsky (2009), he explains that “workers that are infected, poisoned, injured or affected by other harmful factors start to forage and perform other risky tasks at an earlier age than their healthy nest mates.”
Practical effect: You can see how an in-hive epidemic of viruses or nosema can push bees into foraging behavior faster than the broodnest can replace them, which can result in the rapid depopulation of the hive!
This ingenious feedback arrangement allows the colony to best conserve its workforce, yet still be able to take advantage of any sporadic honey flow at the drop of a hat. Indeed, during the flow, the mid-age nectar receivers may flood the broodnest with curing nectar, and effectively shut down the queen’s egglaying (provided that there is not enough drawn comb available directly above the broodnest). This frees up even more nurse bees to take the place of nectar processors that have graduating into foraging! Once the flow is over, the bees can then “dry out” the broodnest, and the queen can get back to laying to replace the often substantial portion of the population that is lost in the frenzy of foraging.
Practical application: Note how the storage of nectar in the broodnest can shut down broodrearing. It makes a big difference exactly where supers of drawn comb or foundation are placed, and how queen excluders are used. Some simple hive manipulations (such as reversing the brood boxes or “checkerboarding”) to create a “break” through the honey band above the brood nest, so as to better allow the nectar transporters access to the empty combs above, can be used to help control swarming, and to produce larger honey crops (see the observations by Walt Wright).
In those areas of Australia with long, intense honeyflows, beekeepers keep the broodnest from becoming plugged by running a single brood chamber below an excluder, and by placing supers of wet drawn comb (“stickies”) directly above the excluder as often as every week. This practice keeps open comb directly above the brood—exactly where the mid-aged bees prefer to place nectar.
A point to note is that BP also affects the foragers—it encourages them to forage for pollen. So during a strong nectar flow, when there is little young brood, there is less incentive for foragers to restock the broodnest with pollen. This can have consequences down the road, when the colony tries to rebuild after the main flow.
Harris (2008a) found that “During the summer, most bees [die] within 5–7 weeks of emergence, so colonies must maintain very high reproductive rates if they are going to produce large colonies that are biologically or economically viable.” The point is, that as soon as broodrearing slows (generally in late July), the colony population can drop rapidly, since workers continue to die at an early age.
Practical application: After a strong honeyflow (especially a pollen-poor one), the colony may be lacking in enough pollen to recover its population by new broodrearing. This can lead to colonies going into fall in weak condition!
One last item of interest: When a colony is starving, there is no reason for young bees to waste time nursing or building comb—the colony needs food, and fast! When emerging bees don’t find enough food in the hive, they tend to simply fast-forward and begin foraging precociously (and preferentially for nectar) (Schultz 1998, Nelson 2007, Marco Antonio 2008).
OK, that’s enough for now—we’ve covered the right side of the model. Next month I will continue with a discussion of the primer pheromones, and the colony reproduction on the left side of the model.
Update Nov 2013 Dr Brian Johnson proposed an excellent overall model of task allocation in the honey bee colony, freely available at:
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2810364/
Johnson, BR (2010) Division of labor in honeybees: form, function, and proximate mechanisms. Behav Ecol Sociobiol 64:305–316.
Acknowledgements
I could not begin to compile the information necessary for my writing without the steady assistance and suggestions by my friend and collaborator Peter Loring Borst, to whom I am again greatly indebted. I also wish to express my gratitude to some of the giants in this field of research who took time to review this article for accuracy: Drs. Gro Amdam, Zachary Huang, and Gene Robinson. I also appreciate the generous correspondence from Lloyd Harris, and Drs. Heather Mattila and Brian Johnson. I would also be greatly remiss not to tip my hat to that great bee behavioralist, Dr. Tom Seeley, who has inspired a generation of bee scientists!
References
Amdam, GV, et al (2004) Hormonal control of the yolk precursor vitellogenin regulates immune function and longevity in honeybees. Experimental Gerontology 39 (2004) 767–773.
Harris, JL (2008a) Development of honey bee colonies initiated from package bees on the northern Great Plains of North America. Journal of Apicultural Research and Bee World 47(2): 141–150.
Higes, M, et al (2008) How natural infection by Nosema ceranae causes honeybee colony collapse. Environmental Microbiology 10(10): 2659–2669.
Johnson, BR (2009) A self-organizing model for task allocation via frequent task quitting and random walks in the honeybee. The American Naturalist 174(4):
Leoncini, I., Le Conte, Y., Costagliola, G., Plettner, E., Toth, A. L., Wang, M., Huang, Z., Bécard, J.-M., Crauser, D., Slessor, K. N. and Robinson, G. E. (2004) Regulation of behavioral maturation by a primer pheromone produced by adult worker honey bees. Proc Natl Acad Sci USA 101: 17559-17564.
Lourenço AP, Martins JR, Bitondi MMG, Simões ZLP.(2009) Trade-off between immune stimulation and expression of storage protein genes. Arch Insect Biochem Physiol. 71(2): 70-87.
Lin HR, Dusset C, Huang ZY (2004) Short-term changes in juvenile hormone titers in honey bee workers due to stress. Apidologie 35: 319–327.
Marco Antonio DS, Guidugli-Lazzarini KR, Nascimento AM, Simões ZLP, Hartfelder K.(2008) RNAi-mediated silencing of vitellogenin gene function turns honeybee (Apis mellifera) workers into extremely precocious foragers. Naturwissenschaften. 95: 953-961.
Pankiw T. (2004) Brood pheromone regulates foraging activity of honey bees (Hymenoptera: Apidae). J Econ Entomol. 97(3):748-51.
Schulz, DJ, ZY Huang, and GE Robinson (1998) Effects of colony food shortage on behavioral development in honey bees. Behav Ecol Sociobiol 42: 295-303.
Tofilski, A (2009) Shorter-lived workers start foraging earlier. Insect. Soc. 56(4): 359-366.
Toth, A.L. et al., 2005. Nutritional status influences socially regulated foraging ontogeny in honey bees. Journal of Experimental Biology 208, 4641–4649.
Wright, W (various) www.beesource.com/point-of-view/walt-wright/
Suggested Review Papers
Johnson, BR (2010) Division of labor in honeybees: form, function, and proximate mechanisms (Review). Behav Ecol Sociobiol 64:305–316. Free download http://berkeley.academia.edu/documents/0065/9441/Johnson_2010a.pdf
Pankiw, T (2004) Cued in: honey bee pheromones as information flow and collective decision-making. Apidologie 35: 217–226
Slessor KN, ML Winston, Y LeConte (2005) Pheromone communication in the honeybee (Apis mellifera L.). J Chem Ecol. 2005;31:2731–2745.
Additional Suggested Reading
Huang, Z and GE Robinson (1992) Honeybee colony integration: Worker-worker interactions mediate hormonally regulated plasticity in division of labor. Proc. Natl. Acad. Sci. 89: 11726-11729.
Huang, Z and GE Robinson (1996) Regulation of honey bee division of labor by colony age demography. Behav Ecol Sociobiol (1996) 39 : 147–158.
Mattila HR, Otis GW (2007a) Dwindling pollen resources trigger the transition to broodless populations of long lived honeybee each autumn. Ecol Entomol 32:496–505.
Mattila HR, Otis GW (2007b) Manipulating pollen supply in honey bee colonies during the fall does not affect the performance of winter bees. Canadian Entomol. 139: 554-563.
Maurizio A (1950) The influence of pollen feeding and brood rearing on the length of life and physiological condition of the honeybee: preliminary report. Bee World 31:9–12
Nelson CM, KE Ihle , MK Fondrk MK, RE Page, Jr, and GV Amdam (2007) The gene vitellogenin has multiple coordinating effects on social organization . PLoS Biol 5(3): e62.


