
The Varroa Problem: Part 5- Striking a Deal with Varroa

February 28, 2017
Contents
Striking a Deal with Varroa. 3
Other mechanisms and traits. 4
The strategy for subduing varroa. 6
The Varroa Problem: Part 5
Striking a Deal with Varroa
First published in ABJ February 2017
Randy Oliver
ScientificBeekeeping.com
Look, varroa is here to stay—we’re never going to defeat the mite. There are no more silver bullets in the developmental pipeline. It’s time to give up on the Band-Aids and flyswatters that we’ve been depending on, and shift to a new paradigm. Let’s help our bees to strike a deal with the mite.
Selective breeding
It’s been clearly demonstrated that natural selection can shift an unmanaged honey bee population to some degree of mite resistance within less than a decade. Unfortunately, the resulting bees may not be such that we’d want to keep. But that’s only because natural selection is a non-directed random process based solely upon chance and reproductive success. Humans, on the other hand, can help to direct the process of evolution, and create breeds of plants or animals with any combination of traits that we choose to select for.
Beekeepers have been selecting for bees with desirable traits since the first clay hives in Mesopotamia. The two traits most consistently chosen for (in most cultures) are gentleness and productivity. In modern times, U.S. breeders have focused upon productivity, and have taken a bee that evolved for occupancy of small tree cavities into Goliaths that build up rapidly in the spring for package production, and then continue to brood up to fill tall stacks of supers.
Practical application: those huge colonies that we are so proud of are also the varroa mites’ dream come true—it’s your largest colonies that tend to crash the hardest.
There’s no free lunch
Unfortunately, there are costs to the breeding of such superbees for economic traits alone—poor colony survivorship without benefit of regular husbandry and mite treatments, increased susceptibility to disease, and a quicker burnout of queens [[1]]. There is also the “genetic cost of domestication”—loss of genetic diversity due to bottlenecking, and the possible accumulation of deleterious mutations [[2]].
Two observations hammer this point home to me: (1) the poor survivorship of our managed bees under feral conditions [[3]], and (2) the incredible difference in health and vigor that I observe in hived Southern California feral bees compared to that of domestic stock in the same yards.
I have absolutely the greatest admiration and respect for our queen breeders and producers. As businessmen they produce what the market demands, so I am not about to criticize them in any way. But I am going to point out a serious flaw in current breeding practices. You see, most commercial queen producers are actually involved in several concurrent breeding programs:
- Selecting for bees that perform well under intensive management and heavy feeding.
- Selecting for varroa mites that are the most virulent and most resistant to miticides.
- Selecting for bee gut and disease-causing bacteria that are resistant to antibiotics.
- Selecting for virus strains best adapted to the above environment.
Take a moment to think about the end result of such linked selection programs…
Yep, self-perpetuating headache, comb and honey contamination, “supermites,” and high rates of colony loss. We can do better.
Practical application: the queen producers respond to market demand. When the market demands mite resistance, I have full faith that they will deliver.
The good news
It’s not that we need to go back to tiny colonies of hot, swarmy bees. I’ve been fortunate to have kept colonies that were not only highly resistant to varroa, but gentle and productive as well. There’s no reason that we can’t have the best of both.
Practical application: we simply need to start prioritizing mite resistance as a selection criterion.
What needs to change
Most of us beekeepers spend our time producing varroa food, with maybe a bit of pollination or honey production thrown in as a sideline. This is because our domestic bee stock provides a veritable no-charge, all-you-can-eat buffet for appreciative mites. Most of the bees sold in the U.S. do virtually nothing to hamper varroa in its nonstop orgy of reproduction.
It’s not that varroa is such a formidable enemy. Indeed, it barely ekes out an existence in colonies of its original host, Apis cerana. In resistant stocks of Apis mellifera the mite doesn’t disappear, but the bees prevent it from increasing to economically-damaging levels.
Striking a Deal with Varroa
Beekeepers rationalize that varroa is a tough, invincible enemy. In actuality, nothing could be further from the truth. Varroa is actually a blind, defenseless parasite, completely dependent upon its senses of smell, touch, and (presumably) taste. And it’s an obligate parasite, able to survive only in the warm, humid environment of the bee hive, and needs to feed frequently on protein-rich bee tissue. The fragile male and immature female mites quickly die outside of the safe confines of a sealed brood cell, and even a mature female can succumb to the bite of a bee. Every step of the mite’s life cycle depends upon the finding of safe locations and the sensing of tiny details in olfactory cues that allow it to synchronize its reproductive timing with its host pupa’s development.
In the distant past, an ancestor of Varroa destructor was somehow able to establish a permanent home in A. cerana colonies. Long-term coevolution led to a relatively stable host/parasite relationship, since it was in the interest of the mite that its host be able to maintain a thriving population. Thus, cerana adapted to cope with the mite, and varroa settled for being able to eke out a tenuous existence in the hive (as a vertically-transmitted minor parasite).
Until recently, I suspected that the suite of coping mechanisms used by cerana against varroa should be the model for mellifera. But it turns out that there are likely an infinite number of ways for bees to manage this parasite. But let’s first look at how cerana and varroa reached a coadaptive “agreement” (as described by Rath in 1999 [[4]], with one critical component only recently elaborated by Page [[5]]).
To start with, let’s be clear who set the rules. Since varroa are like little robotic reproducing automatons, it was up to the bees. So here’s the deal that the bees offered to its unwanted houseguest:
The House Rules
- We don’t like you being here, and we will do everything possible to make your life miserable.
- We will allow you to reproduce only in the drone brood (the carrot), with a few conditions (the stick):
- Our drone prepupae will give off a clear kairomonal signal so that you blind mites will be able to easily differentiate them from worker prepupae [[6]].
- We will keep an opening in the top of the drone capping, and if the drone pupa gives off an odor signal that you have harmed it, we will entomb you and the pupa together into that cell.
- We will limit your reproduction by only rearing drone brood from time to time.
- If you attempt to feed on a worker pupa, it will generally sacrifice itself. If we detect that you are reproducing on a worker pupa, we will uncap the cell and kill you (VSH).
- At every chance, we will groom you off of ourselves and each other, and bite your legs off.
The mite, having no choice in the matter, in turn developed a curved protective “shell” adapted for hugging tight to the curves of a bee’s body. It also developed chemical camouflage by modifying its exoskeleton to quickly absorb the scent of its bee hosts, thus rendering it invisible to the bees in the darkness of the hive. Once a mite scrambles out of reach of an unfortunate new mount, it can escape grooming so long as it stays put [[7]]. This is how mites survive in cerana colonies between bouts of drone rearing—a mite may only get one chance in its lifetime to reproduce.
Practical application: the mite’s Achilles’ heel is that it must shift to younger workers from time to time, otherwise all the mites would eventually be carried out of the hive by the exiting of older workers on their final flight. During this shift to a new mount, mites are exposed to the vigorous auto- and group grooming by the workers.
Thus, life for varroa is not easy in an A. cerana hive. By restricting how often they rear drone brood, cerana can manage to keep the mite population to a tolerable level. The “deal” worked out between cerana and varroa is certainly a workable and successful model, but not the only way to skin the varroa cat.
Other mechanisms and traits
- mellifera, unlike A. cerana, has not (yet) developed the drone brood entombment, worker brood self sacrifice in response to wounding, nor the exclusion of varroa reproduction to drone brood alone. But that hasn’t kept various races and breeds of A. mellifera from developing degrees of resistance to the mite, based upon various combinations of traits [[8]].
Scientific terminology: a phenotypic trait is a distinct variant of an observable and measureable characteristic, such as body color, degree of defensive behavior, or the particular semiochemicals expressed by a larva. Any trait is the result of the expression of the specific alleles (variations) of the gene(s) controlling that trait, in combination with other epigenetic and environmental influences on the expression of that trait (Fig. 1). As an example, vigorous grooming behavior by a worker in response to feeling a mite climbing on its body is a trait.
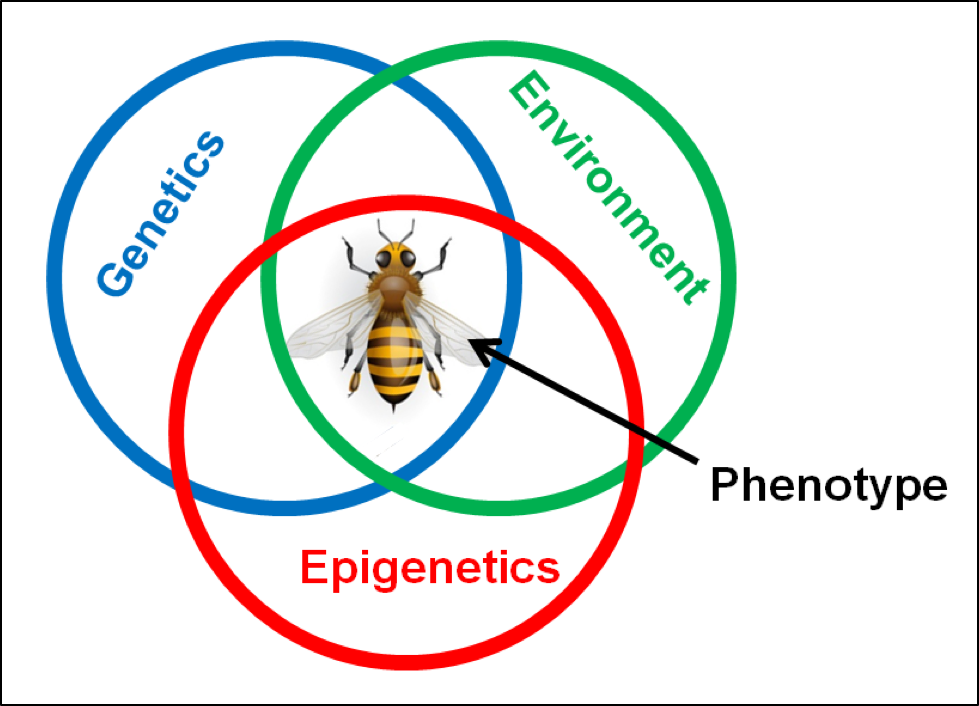
Figure 1. The phenotypic traits (observable variations in form, color, behaviors, or physiology) of a bee are the result of inherited genetics and epigenetics, as well as the regulation of gene expression due to environmental cues or triggers. Many of the traits for varroa resistance may not require novel mutations, but rather simple up- or down-regulation of existing genes. A genetically-diverse population of bees will contain a large “toolkit” of potential traits, due to the presence of a variety of alleles for each gene (which can then be used in various combinations), as well as a diversity of epigenetic expressions.
Practical application: natural selection, due to the degree of randomness and chance involved, might come up with any number of combinations heritable traits that might confer a degree of mite resistance.
Progress can be fast
With strong selective pressure to weed out mite-susceptible bloodlines, a large population of bees can shift to exhibiting a substantial degree of mite resistance surprisingly quickly. I suspect that the skilled queen producers that I know could accelerate the process, if they put in the effort. It’s not that we necessarily need any miraculous new mutations—sexual recombination and epigenetic regulation can effect very rapid heritable changes (Fig. 2).
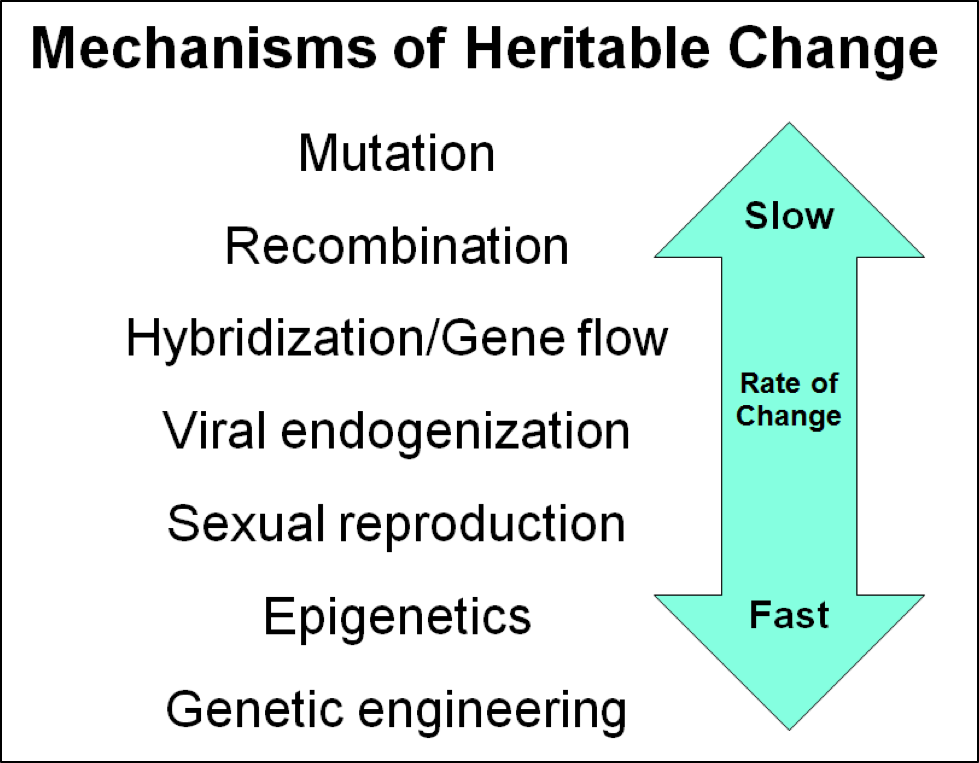
Figure 2. The Earth is not a stable place for life. Some forms of life adapt quickly following volcanic eruptions, asteroid impacts, ice ages, desertification, and the invasion of novel predators, parasites, or food sources (the losers go extinct). There’s no reason to think that our bees can’t come up with a winning hand from their existing deck of cards. But we’ll never know unless we get serious about playing the game.
Knowing your enemy
The military strategist Sun Tsu advises to know your enemy as well as you know yourself. From that knowledge, you can understand your enemy’s strengths, and attack his weaknesses. And this is how we can help our bees to fight varroa—to look for the mite’s Achilles’ heels (it has a slew of weak points). Sun Tsu also points out that one wins battles by having an overall strategy, and then applying specific tactics.
The strategy for subduing varroa
Varroa is in heaven when colonies are engaged in serious broodrearing, especially when there is an abundance of drone brood. During this period of time (the linear buildup phase of a colony) varroa is able to successfully reproduce at a high rate, better than doubling its population each month.
Scientific note: the percent increase in a population over time is termed the population growth rate. Under ideal conditions, that growth rate is exponential, and is quantified as the intrinsic rate of increase. Biologists often break that rate of increase down to a daily value termed “r (day).” The r (day) of varroa in nonresistant European bees during periods of broodrearing is typically around 0.021 (Fig. 3).

Figure 3. So long as brood is continually present (as opposed to the sporadic drone rearing of A. cerana), the mite population in a typical European honey bee hive will increase in an exponential manner [[9]]. As illustrated by the simple graph above, the mite population grows in a typical colony of commercial stock from 100 to over 8000 in seven months. On the other hand, in African bees the population reaches an asymptote at around 1000-3000 mites.
Practical application: the rate of varroa buildup is exponential at all times when brood is abundant (nearly doubling each month)—you just don’t notice it until the late-season decrease in the bee population makes it seem that varroa suddenly exploded. In addition, an influx of mites carried by robbing foragers and drifting bees can boost the varroa population at that time.
Of interest: That burgeoning mite population does not consist entirely of fertile young females. During the season, the proportion of unfertilized, non-reproducing, or plain worn-out mites increases [[10]]. Although they may no longer successfully reproduce when they enter a cell, they still continue to transmit viruses when they feed upon workers or brood.
The above graph looks pretty grim. Luckily there’s another side to the coin—when broodrearing is greatly reduced or absent, the mite population will drop, due to natural death, emigration on exiting bees, and grooming. Thus, an overall strategy for mite resistance would consist of two parts:
- Increasing the death rate of the mite at all times, and
- Decreasing the birth rate of the mites when brood is present.
The bees’ tactics
INCREASING THE DEATH RATE
Grooming and biting: As far as I can tell, most breeds of mite-resistant bees exhibit vigorous grooming or attacking of mites so long as they are moving on a bee’s body [[11]], and some breeds are good at biting and killing them. Thus, the mite death rate can be increased by selecting for grooming/biting behaviors (and perhaps modification of the bees’ mandibles [[12]]). Bees need to be bred first to recognize the presence of a mite, and then for the behavior to grab and kill it. There are also behaviors for self grooming, and doing a “grooming dance” to initiate grooming by nestmates. As mentioned before, a mite residing in a colony of “mite biters” faces a tough choice as the bee that it’s riding on ages—if it doesn’t move to a younger bee from time to time (thus exposing it to grooming), it will eventually be carried out of the hive on its ride’s last flight. The act of interrupting mite reproduction by VSH also increases mite mortality by exposing the foundress to the workers.
A common question: What happens to the foundress mite after the bees chew out the pupa during the process of VSH—can they still enter another cell and reproduce?
Once a foundress mite begins to ovulate within the safe confines of a capped cell, her belly begins to swell [[13]]. If she is chased out of the cell at that time by bees exhibiting VSH behavior, she can no longer fit tight against a bee and stands a greater chance of being caught by the bees. Not only that, but that mite is now reproductively out of synch, and even if she quickly enters another brood cell, she will no longer lay a male egg, thus producing infertile daughters [[14]].
By increasing the mites’ rate of daily mortality, the bees can partially offset mite reproduction when brood is present, reduce the average number of cycles that a mite is able to reproduce, and accelerate the decrease in the mite population during winter. Dr. Greg Hunt has clearly demonstrated that there is great variation among colonies in the rate at which they groom off mites [[15]].
The Primorsky Russian bees exhibit a suite of traits that in total help to keep varroa in check [[16]]. In my area, during the late-summer dearth, they completely cease broodrearing, thus allowing them to groom off the mites that built up during spring and early summer. The colony then later resumes broodrearing during the fall bloom, starting with a much reduced level of mites.
A thought: consider the effect of expanding the trait of altruistic self removal behavior that bees already exhibit when they feel sick [[17]]. If a bee, should it feel a mite gnawing a feeding wound into its abdomen, immediately sacrifice itself by exiting the hive, varroa would no longer be a problem. My point: there are likely plenty of undiscovered traits for mite resistance.
DECREASING THE BIRTH RATE
Now let’s address the varroa birth rate, for which there are a constellation of traits, which in combination, can result in reducing the overall rate of varroa buildup in a colony. I plan to go into these traits in greater deal later on, but let me hit some basics right now.
VSH: In general, mite-resistant bees exhibit a high degree of varroa sensitive hygiene (VSH), which appears be a sensory and behavioral response to olfactory cues produced by injured, sick, or virus-infected prepupae and pupae, and possibly to the presence of reproducing mites (perhaps due to detection of the mite mating pheromone). VSH also results in a larger proportion of the total mites in the hive at any time being in the phoretic state, where they are more susceptible to grooming.
The trait of “bald brood”—in which the bees uncap pupae but do not remove them has yet to be explained—perhaps it confers some benefit against varroa (possibly due to disruption or dehumidification of the mite family within the uncapped cell) (Fig. 4).

Figure 4. It’s possible that the uncapping of cells to create “bald brood” has something to do with mite resistance. At the top of the photo there is a chewed out pupa (VSH), and in the lower right what appears to be an uncapped but dying prepupae.
SMR and semiochemicals: Since varroa is completely dependent upon specific odor cues at every step of its life cycle (especially for the identification of the correct age and type of brood to parasitize, the precise timing to initiate ovulation, to allow a male to identify a female ready to mate, etc. [[18]]), by tweaking those chemical cues slightly, the bees may be able to mess with varroa’s success in reproduction. This tactic goes by the name of suppression of mite reproduction (SMR).
Allow me to clearly state that SMR is a controversial subject among mite-resistance researchers. The main problem is that it is difficult to separate SMR from VSH unless during assessment the cells are protected by screen shortly after capping (in order to prevent removal of infested pupae [[19]]). That said, my impression is that because the mites are so utterly dependent upon those precise odor cues, that this trait has potential for the bees to use to reduce mite reproductive success, an opinion that is bolstered by the strikingly poor reproductive success of varroa in the brood of African and Africanized bees [[20]].
Outside the box thinking: what if bees evolved so that late-stage pupae produced mite mating pheromone (already similar to a bee pheromone) just as the first daughter mites became sexually receptive—would the male mites would then be unable to locate a female?
Postcapping duration: Mike Allsopp [[21]] found that the success of the Cape Bee was partially due to its shorter postcapping duration (average 265 hrs vs. average 282 hrs for European stock), but little progress has been made in the breeding of bees for this trait.
Small cell: Much has been made of the smaller cell size of Africanized bees, but there is little evidence that it substantially contributes to mite resistance—varroa, when it first invaded South Africa, grew rapidly in those naturally small-cell hives. However, there are other mechanical traits, such as certain pupal movements, or the thickening of the area that mites prefer for feeding, that could screw up mite success.
Self-sacrifice of pupae: The pupae of breeds selected for VSH, when they sicken, apparently produce pheromonal signals that signal mid-age bees to remove them. This is only a small step from the “social apoptosis” exhibited by the pupae of A. ceranae, and clearly a trait to look for. This one trait, if we could fix it in a stock, would be a huge step towards solving The Varroa Problem.
Clearly, there are a number of chinks in varroa’s armor. None of the traits above would alone confer mite resistance, but combinations of additive and interactive traits working in concert could certainly do the trick.
Practical application: I’ll get into the specifics more deeply when I get to the modeling of varroa populations later in this series, but as pointed out by Corrêa-Marques [[22]], our goal for a bee stock would be to reduce varroa’s overall “effective reproduction rate”—the number of viable, mated daughters that successfully emerge and survive per each foundress during a mite reproductive cycle under the capping.
Practical question: just how much of a change in only two parameters—grooming and the effective reproduction rate—would it take to produce a mite-resistant bee?
Using models
As suggested by Wilkinson and Smith in 2002 [[23]], modeling can be used to compare the degree to which various biological mechanisms would be expected to decrease peak mite population in a hive. A number of researchers have developed complex models, which have been quite instructive. Unfortunately, those models are generally not very user friendly to Joe Beekeeper. Therefore, I’ve spent a great deal of time in developing a handy mite population model (which I hope to soon have online).
My model allows the user to input the colony broodrearing and growth parameters for your region and operation, as well as the adjustment of multiple variables, including the mean number of daughters per foundress in either worker or drone cells, the mean percent of those daughters that successfully mate and emerge, and the daily mortality rates of mites during the summer and winter. It also shows the effect of mite treatments. As an example, for the simulation below (Fig. 5), I plugged in typical values for an untreated strong hive of commercial bee stock in an area with a 3-month winter brood break.
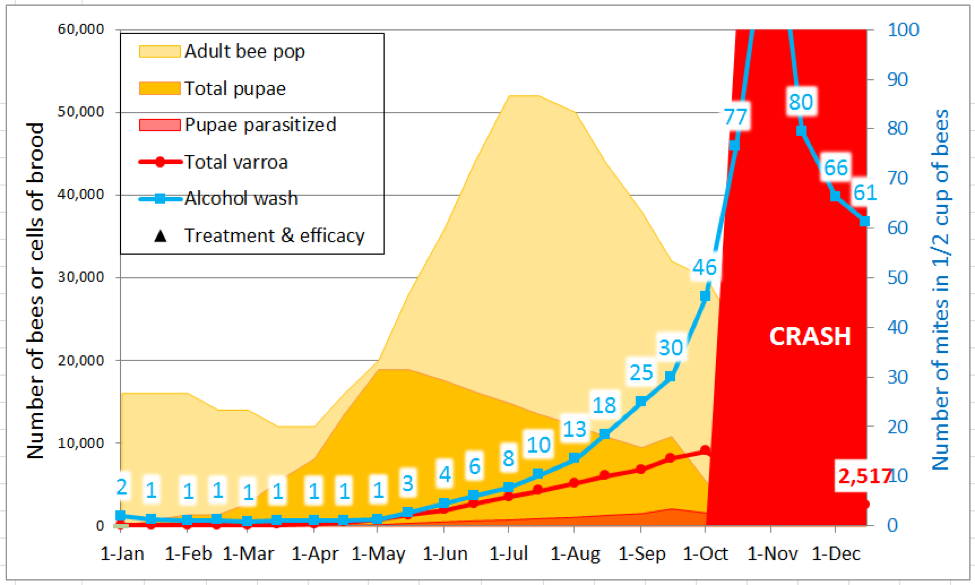
Figure 5. A simulation starting with 100 mites, with minimal mite immigration in late summer, for typical domestic bee stock. During the active broodrearing period of May 1 – July 15, the model calculated an average r (day) of 0.023, with the mite population peaking at 9000 on October 1st. The model arbitrarily indicates colony crash whenever the alcohol wash exceeds 75 mites.
In the above simulation, the rate of increase at either side of peak broodrearing (refer to the orange curve) was less than 0.023, and was positive for only 7½ months (the rest of the year, the r-value was negative, resulting in a drop in the mite population). At colony crash, nearly 50% of pupal cells were invaded by at least one mite. Note that alcohol wash counts wouldn’t have been alarming until July.
In the simulation below (Fig. 6), I left all parameters the same, other than factoring in a mere 20% decrease in the number of daughters per foundress as well as in their success at getting mated (the previous two parameters would be at least partially affected by VSH and SMR), plus 20% increases in the mite mortality rates in summer and winter (likely affected by the grooming/biting traits).

Figure 6. In this simulation, a mere 20% improvement in a few critical traits resulted in an easily manageable mite population at the end of the season (it peaked at under 900 mites in early October). The rate of mite buildup during the May 1 – July 15 period was reduced to an r (day) of only 0.012. For sustainability, the ending mite count of 242 mites could be reduced back to not more than the starting 100 by an oxalic dribble in the fall, or by drone trapping, splitting, or other method in the spring.
Practical application: a mere 20% improvement in only three traits could nearly make varroa a non issue. My point is that we don’t need to make much improvement—there’s already greater than 20% variability in those traits in our existing bee stocks. We just need our large-scale breeders to start selecting for those queens.
Varroa tolerance
I’ve been focusing upon colony resistance to varroa, but there are also mechanisms for tolerance to both the mite and its associated viruses, such as:
Reduction in drone rearing: Although this trait would likely be nonadaptive under natural selection, it could certainly be of benefit to managed bees.
Shorter worker longevity: As pointed out by Martin [[24]], in Africanized bees, varroa populations stabilize at around 1000-3000 mites in a colony, apparently due to the shorter longevity of the workers, coupled with the poor reproductive success of the mites.
Restriction of broodrearing: by starting and ending broodrearing earlier and later in the season, by maintaining smaller broodnests and overall colony size [[25]], or by curtailing broodrearing during pollen dearths (the Russians) or at other times [[26]], bees can reduce varroa buildup.
Swarming or absconding: Sending off a swarm, by my calcs, will reduce the hive’s mite population by roughly 20-25%, and allow the swarm to begin with a much lower overall infestation rate (since the majority of mites are left behind in the brood).
Virus resistance: Most of the above traits would not be desirable for production stock, but there is one more mechanism of resistance that is. As I explained earlier in this series, the problem isn’t really varroa—it’s virus (generally DWV) that kills the colony. Colonies can tolerate extremely high levels of mites if a virus epidemic is avoided. A key point is that when Nature is allowed to do the selection, it may wind up producing colonies that don’t carry much virus load, as in the case of A.m. scutellata and Africanized bees [[27]]. Or it may not, as in the case of the “Bond Bees” in the Gottland experiment, which survive despite DWV [[28]]. In my own operation, as well as in the Southern California feral stocks that I’ve examined, I find colonies that despite having high mite levels, maintain healthy broodnests (Fig. 7).

Figure 7. Some colonies, such as this one of Russian stock, exhibit high mite counts, yet no sign of disease due to virus in the brood. This places me, as a breeder, in a quandary—whether I should use such high-mite queens into my breeding program (I haven’t, but that doesn’t mean that you shouldn’t). We may have more answers once Dr. Stephen Martin finishes viral analysis of the 500 bee samples that volunteers from across the U.S. recently sent at my request.
wrap up
OK, our existing bee population (domestic plus feral) likely contains all the tools necessary for varroa resistance. But relax—you don’t need to know how the bees go about doing the job. We just need to select for bees that somehow figure out how to get the job done. Next (hopefully): Varroa-Resistance Breeding for Dummies.
aknowledgements
I cannot begin to express my gratitude to Dr. Stephen Martin, whose incredible body of work on varroa and DWV has been instrumental to my research and understanding of our nemesis. As I was putting the finishing touches on this article, I was serendipitously fortunate to be assigned to bunk with him during the Bee Audacious conference in California, which allowed us hours of intense discussion (he’s wagered a bottle of whiskey against me being able to breed resistant stock).
I also wish to express my appreciation to the other (mostly European, but some American) varroa researchers who devoted years of incredibly tedious and difficult work in the early days of the mite invasion (the most fruitful period was from the mid ‘80’s through the early 2000’s). I’ve spent this past year immersed in reading their research (thanks to Peter Borst for helping me to locate those papers). That generation of researchers is aging, and what have we got to show for it? We owe it to them to develop mite-resistant stock while they are still alive to see it happen.
notes and citations
(Most of the papers cited below are (with a bit of diligence) available open access on the Web).
[1] I suspect that the longevity of queens is not a chronological measure, but rather has more to do with the cumulative number of eggs laid over lifetime. Queens in short-season smaller colonies may thus live for more years than those that maintain huge broodnests from February almond pollination through the end of October stimulative feeding.
[2] Wang, G-D, et al (2014) Domestication genomics: evidence from animals. Annu. Rev. Anim. Biosci. 2: 65–84. Luckily, in bees the haploidy of the drones tends to purge deleterious mutations.
[3] https://scientificbeekeeping.com/whats-happening-to-the-bees-part-5-is-there-a-difference-between-domesticated-and-feral-bees/
[4] Rath, W (1999) Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 30: 97-110.
[5] Page, P, et al (2016) Social apoptosis in honey bee superorganisms. Nature Scientific Reports 6:27210 DOI: 10.1038/srep27210
[6] The kairomonal signal from cerana worker brood doesn’t initiate ovulation by a foundress mite (SMR).
[7] Its preferred location is with its head wedged beneath the second visible abdominal sternite, a location where the bee can no longer reach the mite with its legs. You can observe the mites on the undersides of bees when they walk on the glass of an observation hive or jar. Refer to:
Rath, W (1999) Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 30(2-3): 97-110.
[8] Well-reviewed for both cerana and mellifera by:
Boecking, O & M Spivak (1999) Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidology 30: 141-158. Open access. Why haven’t we made more progress?
For Africanized bees:
Calderón, RA, et al (2010) Reproductive biology of Varroa destructor in Africanized honey bees (Apis mellifera). Exp Appl Acarol 50:281–297. Open access.
And for survivor mellifera lines by:
Locke, B (2015) Natural Varroa mite-surviving Apis mellifera honeybee populations. Apidologie DOI: 10.1007/s13592-015-0412-8 Open access.
[9] The function is P(t) = P(0) * ert.
[10] Corrêa-Marques, MH, et al (2003) Comparing data on the reproduction of Varroa destructor. Genetics and Molecular Research 2(1): 1-6.
[11] With the highly mite-resistant Cape honey bee being a notable exception.
[12] Peng, Y-S, et al (1987) The resistance mechanism of the Asian honey bee, Apis cerana Fabr, to an ectoparasitic mite, Varroa jacobsoni Oudemans. Journal of Invertebrate Pathology 49: 54-60.
[13] Rath, op cit.
[14] Kirrane, MJ, et al (2011) Asynchronous development of honey bee host and Varroa destructor (Mesostigmata: Varroidae) influences reproductive potential of mites. J. Econ. Entomol. 104(4): 1146-1152. But this was not supported by:
Garrido, C & P Rosencranz (2004) Volatiles of the honey bee larva initiate oogenesis in the parastitic mite Varroa destructor. Chemoecology 14: 193-197.
[15] Andino, GK & GJ Hunt (2011) A scientific note on a new assay to measure honeybee mite-grooming behavior. Apidologie 42:481–484.
[16] Rinderer, TE,., et al. (2001) Resistance to the parasitic mite Varroa destructor in honey bees from far-eastern Russia. Apidologie 32(4): 381–394.
[17] Rueppell, O, et al (2010) Altruistic self-removal of health-compromised honey bee workers from their hive. J. Evol. Biol. 23: 1538-1546.
[18] I’m curious as to whether the prepupal cues also affect whether or not a foundress mite first lays a male egg, since in some breeds of bees, there appears to be less success in the mites doing so.
[19] Unfortunately, few researchers have bothered to do this critical step.
[20] Strauss, U, et al (2015) Resistance rather than tolerance explains survival of savannah honeybees (Apis mellifera scutellata) to infestation by the parasitic mite Varroa destructor. Parasitology 143: 374–387.
[21] Allsopp, M (2006) Analysis of Varroa destructor infestation of Southern African honeybee populations. M.S. Thesis, Univ. of Pretoria. http://upetd.up.ac.za/thesis/available/etd-08082007-153050/unrestricted/dissertation.pdf A must read for anyone interested in breeding bees for varroa resistance.
[22] Corrêa-Marques, op. cit.
[23] Wilkinson, D & GC Smith (2002) A model of the mite parasite, Varroa destructor, on honeybees (Apis mellifera) to investigate parameters important to mite population growth. Ecol Modelling 148: 263 –27.
[24] Martin, SJ & LM Medina (2004) Africanized honeybees have unique tolerance to Varroa mites. TRENDS in Parasitology 20(3): 112-114.
[25] Loftus JC, ML Smith, TD Seeley (2016) How honey bee colonies survive in the wild: Testing the importance of small nests and frequent swarming. PLoS ONE 11(3): e0150362. doi:10.1371/journal.pone.0150362
[26] I continue to hear reports of ”survivor” colonies in coastal Northern California taking a brood break during summer, despite the availability of pollen.
[27] Pers. comms. Dr. Mike Allsopp & Dr. Stephen Martin, and
Mendoza,M, et al (2014) Nosema ceranae and RNA viruses in European and Africanized honeybee colonies (Apis mellifera) in Uruguay. Apidologie 45 (2): 224-234.
- cerana also appears to be more resistant to viruses than European stock.
[28] Locke B, et al (2014) Increased tolerance and resistance to virus Infections: A possible factor in the survival of Varroa destructor-resistant honey bees (Apis mellifera). PLoS ONE 9(6): e99998. doi:10.1371/journal.pone.0099998


