
The Varroa Problem: Part 7- Walking the Walk

July 28, 2017
The Varroa Problem: Part 7
Walking the Walk
Randy Oliver
ScientificBeekeeping.com
First published in ABJ May 2017
I’m not one to tell any beekeeper what they “should” be doing—it’s up to nature, the market, personal preference, and history to determine what works. In my last two articles, I’ve discussed ways to go about breeding for mite resistance. Now it’s time for me to walk the walk.
Although I hardly qualify as a queen producer, I’ve bred all the queens for my own operation for decades (I currently mate out around 2500 queens a season). And although my selective breeding program would fully qualify as being half-assed, over the years I’ve found it to be surprisingly easy to breed for color, gentleness, productivity, and resistance to AFB, chalkbrood, and tracheal mite.
After my successes at breeding what I at the time considered to be “superbees,” I wasn’t concerned about the arrival of varroa in my operation (1993), figuring that I’d soon be showing off another notch in my belt. Boy was I wrong—varroa wiped me out, and although I was able to give up dependence upon synthetic miticides some 16 years ago, I still depend upon treatments to keep my bees alive.
This is not for lack of effort to breed a better bee. I’ve got nothing to hide as I walk the walk myself in attempting to breed mite resistance into my stock, and perhaps by relating what I’ve done, I might save others from repeating my mistakes, as well as sharing what I’m trying now, and, more importantly, the biological reasons for doing so.
Frankenbees
I first attempted to breed mite resistance based upon wishful thinking rather than biology. I bought all kinds of (often expensive) breeder queens from others who claimed mite resistance, with the grand vision that I’d just put them all in the same yard, let the drones mix up their genes, and abracadabra—pure magic would create The Perfect Bee in my very own yard!
In retrospect, it’s embarrassing that I could have been so naïve. That was like dreaming that I could disassemble a Ferrari, a Porsche, and a Ford, and then randomly reassemble their parts to create the ultimate racing machine. What I got instead was a bunch of worthless Frankenbees. Look, each of those cars was the result of many years of evolutionary trial and error, culminating in vehicles with perfectly-tuned interacting systems. Change one part randomly, and you wind up with junk. It’s generally the same with bees.
People often misunderstand the concept of “hybrid vigor,” thinking that any cross will be better than either parental type. Hybrid vigor (heterosis) may indeed occur when one crosses two artificially inbred strains of domesticated plants or animals (thus returning closer to the wild type), but crossing two wild types more often results in less fit hybrids. This is due to messing up the fine-tuning achieved by generations of natural selection acting upon the genes and epigenetic regulation so that they work well together as a system [[1]].
Allow me to give a simple example. Suppose that there are two regionally-adapted breeding populations of bees, different enough to be classified as separate races—each specialized for optimal fitness in their respective environments (Fig. 1).
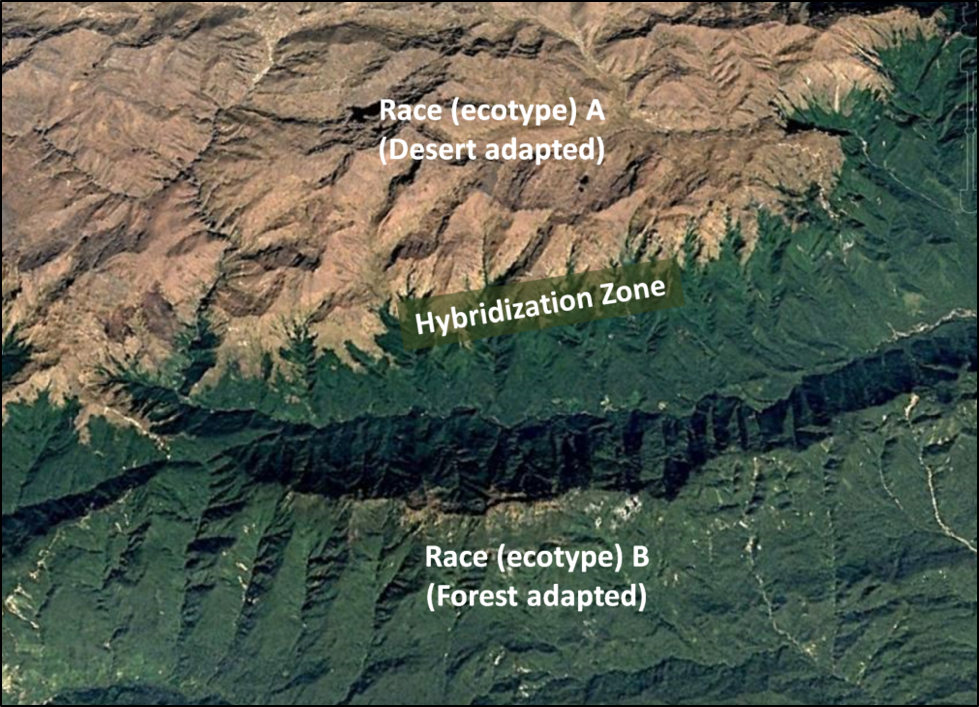
Figure 1. Two separated and regionally-specialized breeding populations (ecotypes) of bees may each be well adapted for their particular environments. But since all races of Apis mellifera are able to interbreed, a continuum of hybridization inevitably occurs between what taxonomists arbitrarily classify as separate “races” [[2]].
In between the ranges of the two races would be a “tension zone” of hybrids, with some degree of gene flow between the races [[3]],) but with the parental populations nonetheless retaining their genetic integrity. If any of those hybrids exhibited greater fitness in either of the parental races’ environments, the hybrid would quickly outcompete and displace the parental type, or it might develop into a race of its own. My point is that we generally don’t see bee races being rapidly replaced by hybrids resulting from human introductions, strongly suggesting that hybrids typically exhibit a lesser degree of “fitness” in that environment than does the naturally-evolved resident ecotype [[4]].
An exception and example: the hybrid Africanized bee clearly exhibited greater fitness than did the preexisting (but artificially introduced, and poorly adapted) European races in South and Central American tropical and subtropical regions. However, note that despite its rapid expansion, the hybrid has not displaced European types above about 30° latitude [[5]], suggesting that the Europeans are better fitted to those ecotypes (refer back to the ecoregion map in my previous article). Note also that the slow but steady flow of alleles and mutations between bee races allows for better evolutionary adaptation, even if you don’t notice a difference in how the bees look or behave.
This does not mean that a true-breeding hybrid miracle couldn’t occur in your breeding program, but the odds of it happening in the short term are not in your favor. Evolutionarily, time is not on the bee breeder’s side, since we typically only have one generation per year to select from, and it may take quite a few generations for a multi-hybrid breeding population to work out the bugs.
My Return to Basic Breeding
In my case, I decided to return to my old ways, and to work with what I already had—locally-adapted stock selected for characteristics that I found desirable. To that, I brought in the occasional instrumentally-inseminated VSH, Russian, and a few other promising breeder queens to produce drone mother colonies.
Practical application: by adding some of those drone mothers to my mating yards, I’d “offer” the new alleles (and genetic and epigenetic combinations) to my own breeding population without necessarily messing up the fine tuning that population already had going. If the new combinations proved to be adaptive, they’d make it into future generations; if not, they’d be bred out, since I wouldn’t select queens from second-rate colonies as breeders.
Then what I’ve done for the past 15 years is to mark potential breeders throughout the season, and to make my final selections each spring after almond bloom. By that time, the colonies had proven their mettle by making honey over the summer, then wintering successfully, and then building up early for almonds (all traits that make me money). I’d only choose the strongest colonies in March (having bees wall to wall), having put on honey in the almonds (as evidenced by weight gain and white wax), and exhibiting gentleness and solid brood patterns. Lastly, I’d perform an alcohol wash to see how much varroa had built up since the fall treatment. There was always enough colony-to-colony variation in mite counts to lead me to believe that this selection process was adequate, and over the years our cutoff for allowable mite counts for our breeders dropped from 4, to 3, to 2, to 1, to mostly zeroes. I’d then choose a minimum of 25 queens each year to use as breeders, in order to maintain some degree of genetic diversity (this may have been a well-intentioned mistake).
Practical application and reality check: although I could see what appeared to be progress towards mite resistance, when I then ran those breeders without treatment for their second season, nearly every one would allow the damn mites to build up to levels requiring treatment, year after frustrating year. That’s why I decided to step up my game, and go for the more formal selection process that I described earlier in this series.
It’s all about the team
Keep in mind that the queen herself can’t directly contribute that much to her colony’s performance, other than by laying a lot of eggs and producing the pheromones necessary for good colony morale. It’s up to her daughters to do the work of rearing that brood, foraging and honey processing, staying calm on the combs, and resisting mites.
A good baseball team consists of players each with specialties—you need a pitcher, a catcher, outfielders who can catch and throw, batters who can hit home runs (or be good runners if they can’t). A team of only batters who couldn’t throw or catch wouldn’t go far. It’s the same with bees. A queen mates with a diversity of drones, each of which then fathers a patriline of sister daughters. A queen that mated with twenty drones will head a colony that consists of twenty patrilines of workers—each patriline with different traits (analogous to each of the players on a baseball team). If the traits of all the players complement each other, then you’ve got a winning team or colony.
The beauty of the honey bee mating system is that in a natural situation, each queen is most likely to mate with the best drones from a cross section of all the best colonies in the vicinity. It seems to me that the honey bee reproductive system has evolved for each breeding population to be able to recover from decimation events, such as forest fire, drought, a severe winter, or the invasion of a new pathogen or predator. The few surviving colonies, due to the genetic diversity held in the spermatheca of the queens, allows for the recovery of much of that diversity as that population recovers, provided that those surviving colonies manage to swarm a number of times before the original queen dies (hence the high swarming rate of wild-type colonies).
Honey bees do everything in their power to avoid inbreeding. Their system of sex determination prevents incestuous breeding, since the female offspring of such matings are non-viable (they become diploid drones, which are generally removed by the nurses). Thus, when we attempt to bottleneck a bee population by selective breeding, we are forced to work against bee behavior. As I’ve stated before, unless we use instrumental insemination, the best that we can do is to breed a deme—a subpopulation of related bees, but with a requisite amount of genetic diversity to maintain good brood viability.
Practical application: unlike as with lab mice or livestock, it’s impossible to maintain a clonal (genetically identical) population of bees (since there would be only a single sex allele). Page and Laidlaw [[6]] calculated that the minimum number of reproducing breeders required for a sustainable “closed” breeding population is in the order of 35-50 each year.
Honey bees like to mix it up, and then filter
So let’s return to the genetic “magic” that might happen in your breeding program. Nature is generally pretty conservative genetically—we humans have roughly 44% of our genes in common with the honey bee [[7]]. But since the background mutation rate of genes is rather low, most species adopted sexual reproduction in order to allow for new combinations of heritable genetics in each generation, in order to allow for evolutionary adaptation to changing environments.
Honey bees take this “shuffling up” of genes to the extreme—a queen may mate with as many as 40 drones, meaning that any of her daughter queens will be likely to have been sired by a different father. Let’s say that you’ve got the dream colony—the success of that colony was due to the teamwork of say 20 different patrilines of daughters working as a team.
Practical application: you graft a bunch of daughters “from that dream queen.” Each of those daughters gets only half of her genes from the mother, and the other half from only one of the drones that the mother queen had mated with (so each virgin may carry only slightly more than 1/2 of the total alleles that contributed to the performance of the originally colony (any of which may have been critical for the success of the “team”).
You then mate that virgin out to a different population of drones. The workers of the resulting colony could now carry as little as 1/4th of the allelic genome of the original dream colony, and no telling whether the 3/4ths of the genes of the worker force consisting of those patrilines will work well together. And it’s the same for every daughter that you raise from that original breeder queen—each daughter colony will be genetically different. It’s little wonder why it is so difficult to replicate a “dream colony.”
Not only that, but in the process of meiosis that creates germ cells (eggs), a phenomenon called recombination occurs. When a queen splits her paired chromosomes in order to create haploid egg cells, some of the genes in each DNA strand (one coming from her father, one from her mother) swap places, resulting in novel combinations of alleles and epigenetics. Honey bees do this at a much greater rate than most other species—about 15 times as frequently as in humans [[8]].
OK, so it’s pretty obvious that honey bees really like to mix up their genes each generation, which allows for rapid adaptation to changing environments, but what about all the new combinations that don’t work well together? How do they purge deleterious mutations and combinations of alleles? Well, they’ve got a trick up their sleeve—a filtering/purifying mechanism.
It’s likely that every beekeeper reading these words has at least one genetic defect—some of you may indeed suffer from what we call “hereditary diseases” or “genetic disorders”. Serious genetic defects are often “masked” by the presence of a “good” copy of that gene inherited from your other parent [[9]]. Drone bees don’t have that luxury—since drones are haploid (only one copy of each gene) their genetics need to work perfectly if they are to survive and successfully mate with a queen.
Practical application: bees mix up their genes each generation, and then filter the new combinations through the drones (and virgin queens) to weed out those that don’t work. This out mating and mixing of genes plays hell on the queen breeder who purchases an instrumentally-inseminated queen, and then attempts to lock those genes into his breeding population. Unfortunately, this has left a bad taste in the mouths of many who tried to propagate mite-resistant stock.
I’ve been lucky to have seen a few colonies over the years that could hold varroa in check (mostly with VSH heritage). But I’ve never been diligent enough to lock that ability in past the second generation, and in each case watched that desirable trait disappear. But the fault was clearly mine, and I realized that it was time to step up my game and get serious about selective breeding.
The opportunity to test my mettle came two summers ago, when I needed to identify 70 hives with high mite counts for a late-summer test of miticides (you need high starting counts to obtain good efficacy calcs). We wound up needing to mite wash over 200 hives in order to find the required number of high-mite colonies. What caught our attention was one colony that had a mite count of zero—10 months after its last treatment. We tracked her for the rest of the season, and into the next spring, and never washed a single mite. She earned the nickname Queen Zero.
Queen zero
I had no idea how Queen Zero’s colony managed to keep the mite level so low, and didn’t want to disturb her to investigate, since for my breeding purposes it didn’t really matter. She was a large, healthy queen, with a nice brood pattern, and her colony was gentle and productive. The question was, was the apparent mite resistance of her colony heritable?
The only way to find out was to rear a bunch of daughters from her. So I grafted about 500 queen cells from her larvae (Fig. 2), and successfully mated out about 400 daughters in my various yards.

Figure 2. Ian and I grafting daughters of Queen Zero in a dark room with headlamps (I brightened the photo so that you could see us). I really don’t know why more beekeepers don’t raise their own queens—it’s one of the most gratifying and cost effective things that we do.
Come July, it was time to start hitting the mites with a knockback treatment. We were crazy busy at that time of season, so I asked Eric and Ian to perform mite washes in each yard of Queen Zero’s daughters until they had identified at least two hives with mite counts not over 1, and then not to apply mite treatments to those hives. This typically took them washing about a half dozen hives (mite counts averaged around 5 for most hives at that time of year). In retrospect, I wish that we’d sampled every one of the 400 daughter hives, in order to have identified even more potential breeders. We then tracked those hives for the rest of the season, and marked any that still had mite counts of 1 or less in November (untreated hives in my area would typically have mite counts at that time of season of 30 or more, or be dead).
Earlier in the season, I’d moved the Queen Zero hive home, and placed her in a test yard chock full of colonies with mite counts averaging around 50-60. There was so much mite drift that Queen Zero’s count went up to 14, so I gave her hive a light oxalic dribble, and finally treated all the surrounding test hives. Her count eventually, and spontaneously, went down to 4 by fall.
Practical application: as noted by Dr. Tom Rinderer during his evaluation of Russian stock for mite resistance [[10]], mingling of mite-resistant hives in with infested hives in late summer can triple the mite count of the resistant bees due to the drifting of mites. Keep this in mind when evaluating your potential breeders—place them all in the same yard.
So, how did our selected Queen Zero daughters fare? We mite washed them all this week—counts ranged from 1 to 10, and Queen Zero was up to 7. I was disappointed by the higher counts (hoping for more zeroes), but none of the colonies had been treated in 11 months, and Queen Zero for nearly 3 years. My mistake of not identifying more potential breeders in July now comes back to bite me—we’ve only got a handful of breeders with really low mite counts—my quandary now is to decide how few queens I’m willing to graft from this season (my fear is too much genetic bottlenecking, and loss of sex alleles).
Practical application: those of you who live in the North, where your colonies get an extended brood break in the winter, have no idea how much more difficult mite management is in a subtropical region. Your alcohol wash counts may increase by perhaps 15x over the course of the season, whereas ours might increase by 50x, with colonies crashing in September (alcohol washes reflect roughly a third of the actual buildup of the mite population). Take home message—it takes a much greater degree of mite resistance for a colony to hold its own in areas not enjoying the benefit of an extended winter brood break.
Recapturing the Magic
I was now convinced that Queen Zero’s low mite counts were not merely a fluke, but an actual heritable trait. Apparently that trait was mostly (if not completely) due to her genetics (as opposed to the genes of the drones with which she had mated), since a good percentage of her crossbred daughters appeared to have inherited it. Her genes had come half from her mother and half from a single drone. But any of her daughters would carry only half the genes of that magical combination. My challenge would be to attempt to recapture that original genetic combination (including the genes of the drones with which Queen Zero had mated) that had proved so successful against the mite.
So here’s the problem. Instrumental insemination would be of little help in recovering the genes of the drones that sired Queen Zero’s workers [[11]]. The only way to recover those genetics would be from her daughters. Look, I make no claim of being any sort of geneticist or super queen breeder, so let me share my thinking out loud…O.K., I just deleted several paragraphs of explanation, realizing that a picture would be worth at least a thousand words (Fig. 3):
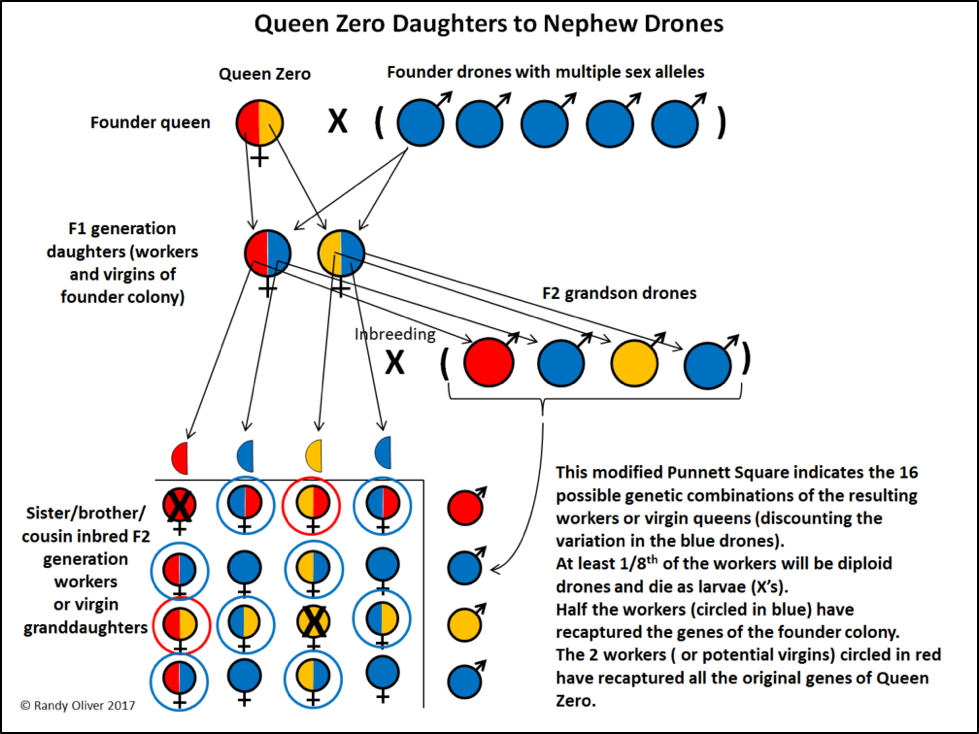
Figure 3. A pedigree schematic of the expected result of some serious inbreeding—crossing her daughters back to the drones of sister colonies started last season. I could recapture a lot of the genetics of Queen Zero herself (as indicated by red and orange), but at the expense of poor brood patterns—on average, at least an eighth of the worker eggs laid by those queens would become diploid drones due to having identical sex alleles.
OK, so what if I eased up a bit on the inbreeding? In Figure 4 I show what would happen if instead of grafting daughters from Queen Zero, I grafted granddaughters, and then mated them out as above.

Figure 4. By mating F2 virgins (Queen Zero’s granddaughters) to drones from a number of sister/cousin colonies, I can still recapture a good deal of the genetics of the original Queen Zero colony, but at less expense (an expected minimum of 6% non-viable brood). I’m intentionally ignoring any problem with sex alleles from the blue or white drones (assuming adequate diversity), but in reality there would likely be some additional non-viability due to duplicate alleles.
Luckily, I now have about 400 marked hives to play with, all headed by daughters of Queen Zero, as well as several hundred unrelated colonies (which likely carry the blue and white genes indicated above). So I now have four main options for breeding, all of which I plan to use:
- I’ll graft some daughters from Queen Zero, and mate them in yards stocked with daughter colonies from last year. This will result in highly-inbred colonies as shown in Fig. 3, but may provide some really good breeding stock for next season.
- I’ll graft virgins from mite-resistant Queen Zero daughter colonies, and mate them as above, to obtain the results shown in Fig. 4).
- I’ll outbreed some Queen Zero daughters and granddaughters by mating them in yards of unrelated drones.
- And I’ll sorta “top cross” some unrelated virgins (from colonies with apparent mite resistance) by mating them in yards filled with drones from F1 daughters of Queen Zero (the drones will be her grandsons). This top crossing is described by Page, Laidlaw, and Ericson [[12]]:
In honeybee breeding, this is equivalent to selecting a single breeder queen (top-cross parent) to contribute all the drones (or a significant proportion) needed to inseminate the virgin queens of a closed population…This is a valuable method for rapidly increasing the frequency of desirable characteristics within breeding populations.
Practical application: let’s say that you wanted to introduce VSH genetics to your own breeding population, but wanted to maintain the traits that you like about your own stock. You could purchase some purebred instrumentally-inseminated VSH breeders. All the drones produced by those breeders would carry VSH genes, and could be used to top cross your existing lines of queens, and work the highly-desirable VSH trait into your queenlines.
I’m going to use the above strategies to severely bottleneck the genetics of my breeding population this generation, and shift it predominately to those of the Queen Zero colony. What, you say, won’t that result in too much inbreeding? After developing the schematics above, I have reason to believe that it won’t be too serious, and will attempt to subsequently rectify that problem by selecting only breeders with solid brood patterns.
Practical application: one method to maintain a diversity of sex alleles in a breeding population is to select only breeders having solid brood patterns—which indicates that the queen carries a diversity of sex alleles other than her own.
A closed population
Ideally, one would practice selective breeding in a “closed population,” which would require an island or a completely isolated mating yard. But I’m happy to have some introgression of “outside” alleles, either from locally-adapted survivor ferals, or from introduced drone mothers (such as VSH breeders). However, the genetics of my bees dominate the landscape, since I provide the majority of nucs and queen cells to the local beekeeping community. I’m also able to completely flood groups of mating yards with my own drones, by making up my nucs from chosen colonies chock full of drones after returning from almonds.
Practical application: unless you are really large scale, there’s slim chance of narrowing your pool of sex alleles too much, unless you only graft off a single queen line for more than one year in a row.
Walking the Walk
I’ll be the first to admit that I’d be a poor choice as a good example of a professional beekeeper. My beekeeping suffers from all my distractions—my experiments and field trials, my literature research, and the number of hours that I spend at writing and speaking engagements. But I do what I most love, and am able to squeak out a living at it.
We beekeepers need to move beyond varroa, and turn varroa management over to our bees. Breeding for mite resistance is indisputably the long term solution to The Varroa Problem. My half-assed breeding efforts to date have shown some success, but I’m as yet unable to dispense with mite management. It’s clearly time to step up my game.
There are others successfully keeping bees without needing mite treatments, and I want to be there too (but without going through the pain and cost of the Bond method). Perhaps by sharing my trials and tribulations in attempting to breed for mite resistance, I can further our collective progress.
Notes and citations
[1] Just to refresh terms—the bees all carry the same genes, but those genes may occur in many slightly different forms called alleles. It is allelic variation, as well as the genetic and epigenetic regulation of the expression of those genes, that causes differences in the phenotypes (the morphology, physiology, and behavior of the individual bee and its colony).
[2] The concept of species, subspecies, and divergent selection is well discussed by James Mallet in:
Mallet, J (2008) Hybridization, ecological races and the nature of species: empirical evidence for the ease of speciation. Philos Trans R Soc Lond B Biol Sci. 363(1506): 2971–2986. Open access.
Mallet (as did Darwin) points out that the concept of “species” is merely a convention useful in communication among biologists. What we call species and what we call populations actually blend imperceptibly into one another, with no clear natural dividing line.
[3] Well reviewed by Byatt MA, et al (2015) The genetic consequences of the anthropogenic movement of social bees. Insectes Sociaux 63:15–24.
[4] Coroian CO, et al (2014) Climate rather than geography separates two European honeybee subspecies. Mol Ecol 23:2353–2361.
[5] Nor 30° south latitude in South America.
[6] Page, RE Jr & HH Laidlaw Jr (1985) Closed population honeybee breeding. Bee World, 66(2): 63-72.
[7] http://ngm.nationalgeographic.com/2013/07/125-explore/shared-genes
[8] Wilfert, L, et al (2007) Variation in genomic recombination rates among animal taxa and the case of social insects. Heredity 98: 189–197.
[9] I’m really simplifying things here. Sometimes, as in the textbook case of sickle cell anemia, a recessive deleterious gene can confer fitness benefits.
[10] Rinderer, TE, L de Guzman, C Harper (2004) The effects of co-mingled Russian and Italian honey bee stocks and sunny or shaded apiaries on Varroa mite infestation level, worker bee population and honey production. Am. Bee J. 144: 481–485.
[11] Since those genes could only be directly recovered from the spermatozoa in her spermatheca.
[12] Page, RE Jr., HH Laidlaw Jr. & EH Erickson Jr. (1985) Closed population honeybee breeding 4. The distribution of sex alleles with top crossing. Journal of Apicultural Research 24(1): 38-42.


