
Understanding Colony Buildup and Decline: Part 11 – Varroa and Late Season Collapse

January 29, 2017
Understanding Colony Buildup and Decline: Part 11
Varroa and Late Season Collapse
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ in April 2016
I’m writing this article shortly after moving my hives to almond pollination. The supply of strong, healthy hives was short this season, but that came as no surprise. I’d heard reports from all over the country last fall that varroa had gained the upper hand. And when that happens, we can expect elevated rates of winter losses.
When varroa first arrived in my apiaries in 1993, I actually looked forward to the challenge. After having successfully bred bees to be resistant to AFB, chalkbrood, and tracheal mite, I was cocksure that varroa wouldn’t be any big deal. Boy was I wrong! In a couple of years nearly all my 250 hives were dead. After nearly thirty years of happy beekeeping, I actually walked away from my deadouts in disgust, frustrated with my feelings of failure and helplessness against this devastating parasite.
The next year my buddy Skip Landon told me about the magic of Apistan strips, and encouraged me to not give up. So I spent the entire summer scraping wax moth out of the abandoned hives and restocking them with bees. To my great relief, beekeeping was again possible—stick in a couple of the magic strips each fall and you’d satisfyingly cover the bottom board with dead mites; the colonies were then good to go until next fall. I even heard a state extension agent tell an audience that varroa was the best thing to happen to professional beekeepers—it reduced our competition from the “bee havers” and feral colonies for nectar sources [1].
I had a great year the next season, but was about to learn a few important lessons:
Lesson #1: Mite immigration from collapsing colonies. I was enjoying my success at having conquered varroa. The next year I treated my hives as per recommendation in early fall and cleaned them of mites. But what I didn’t account for were the feral hives that now had been exposed to varroa for a couple o’ three years. The ferals were now chock full of mites (this was before Deformed Wing Virus gained traction), and that autumn they crashed en masse. Due to drift and robbing, huge numbers of mites reinvaded my treated hives and crashed them in turn (this was the third decimation event of my beekeeping career—first by tracheal mite, then the first varroa invasion, and now from the collapsing ferals). Lesson hard learned: no matter how well you manage varroa in your own hives, they can still be overwhelmed by reinvasion from surrounding hives.
Lesson #2: Viruses evolve. In the first years of beekeeping with varroa, we wouldn’t see bees with signs of Deformed Wing Virus (DWV) until varroa levels were sky high. But that soon changed as natural selection favored strains of the virus that were (1) better adapted to transmission by varroa and (2) were more virulent [2]. No longer could we allow varroa levels to climb as high as we could during the first few years—mite levels that were acceptable in the year 2000 [3] would spell death to a colony today. And I now rarely see bees with deformed wings until long after DWV has already gone epidemic in the brood.
Lesson #3: Varroa evolves too. The bee industry (unlike other ag sectors) has done a poor job at supporting the breeding of bees resistant to our worst pest [4]. But we’ve proven again and again how quickly we can breed for mites resistant to miticides. We bred fluvalinate-resistant mites in about six years; coumaphos-resistant mites in three years; and now there are indications that mites are becoming resistant to amitraz in many areas (Fig. 1).
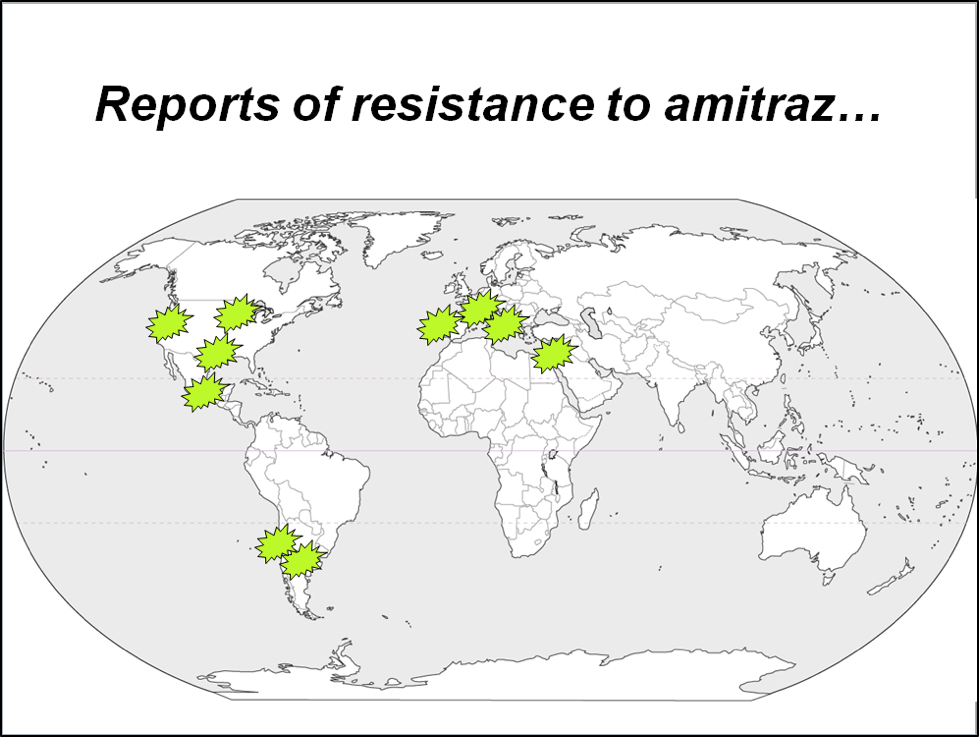
Figure 1. I’ve placed green stars in those areas where amitraz has either anecdotally or scientifically been confirmed as losing efficacy against varroa. Beekeepers, by not practicing “pest resistance management,” keep shooting themselves in the foot.
Our commercial industry has enjoyed a long and successful run with off-label amitraz [5]. Unfortunately, it’s been used by many without rotation, and at far higher doses than necessary (the best use of amitraz would be as a low-dose behavioral disruptor of the mite, rather than for rapid knock down). The time-release Apivar strips can still be effective if used once a year in the springtime, but other mite control measures should then be used for the rest of the season, in order to delay the further development of resistant mites.
Unfortunately, there are few new “silver bullets” in the development pipeline, and I fully expect that our industry is about to enter a new era in mite management—one that is not as dependent upon synthetic miticides. That means that it is increasingly important for beekeepers to better understand the biology of bees and varroa.
Deadly Combinations
It’s understandable that beekeepers want to blame the death of their colonies upon anything other than their own negligence, but the sad fact is that inadequate varroa management is still our number one problem. That said, the progression of colony morbidity is often (to use a term favored by researchers) “multifactorial.” Nutrition, pesticides, other parasites, management, and weather may all enter into the equation, as do colony population dynamics. So let’s take a look at what’s happening in the hive during the late summer decline. Let me be very clear that my experience is mainly with beekeeping in the California foothills, where the honeyflow typically ends by the 4th of July, and pollen becomes scarce until the autumn rains (hopefully) arrive. Pollen availability varies greatly by location—this discussion relates to those times of the summer during which there is a pollen dearth. So you can shift the dates in the chart below to match what occurs in your area (Fig. 2).
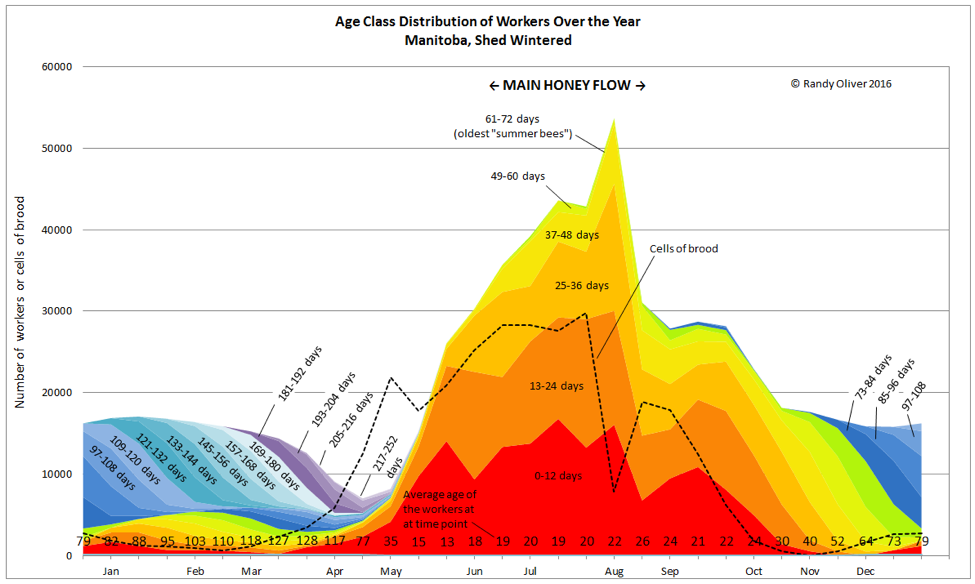
Figure 2. Let’s focus upon the period from late August through early October [[i]]. Note the decline in the amount of broodrearing (dotted line) as a result of reduced pollen availability [[ii]]. This is predictably followed by a drop in the adult bee population. Note also the “aging” of the worker population (bottom line of figures).
[i] Based upon Lloyd Harris’ data cited previously in this series.
[ii] I checked two studies to confirm that pollen income declines in Canadian apiaries at that time of year:
Mattila, HR & GW Otis (2007) Dwindling pollen resources trigger the transition to broodless populations of long-lived honeybees each autumn. Ecological Entomology 32: 496–505.
Nelson, DL, et al (1987) The effect of continuous pollen trapping on sealed brood, honey production and gross income in Northern Alberta. ABJ Aug 1987: 648-650.
The main driver of colony population decline is the reduction in incoming nutritious pollen. From the Spring Turnover on, the colony had exuberantly been rearing brood, which allowed it to maintain a youthful, vigorous, expanding, and resilient workforce. But this comes to a halt as pollen becomes scarce—as the recruitment of new workers plummets, the population dwindles and ages, and may suffer from nutritional stress. From Spring Turnover until the end of the main flow, colonies generally have few health issues—they can usually shake off EFB and nosema—and are trouble free. But things now change.
Practical application: what occurs to me is that before varroa we likely just didn’t notice how close colonies got to the edge of failure during late summer. Varroa (and perhaps Nosema ceranae) changed all that—as my friend Dave Mendes observed, bees are just more “fragile” these days.
I’ve kept bees in the Sierra Foothills since well before varroa. Before the mite, colonies used to survive the summer and winter just fine, despite the lack of floral resources after mid July. But come varroa, I had a helluva time keeping colonies alive unless I moved them to out-of-state pasture for the summer. Finally, tiring of all the driving, I found that by feeding pollen sub I could keep my hives near home for the summer. What I hadn’t grasped is how close colonies used to come to the tip point during our summer dearth—all it now takes is a moderate varroa/virus infestation to push them over the edge.
Side note: Australia offers us a “control group” to which we can compare our beekeeping problems, since varroa has not yet arrived there, and many operations have minimal exposure to pesticides, since their bees forage on wildlands. Yet in the Jan/Feb issue of Australia’s Honeybee News, Dr. Doug Somerville addresses reports of poorly-performing or dying colonies. He concludes that “the general thinking is that, in most cases, poor performing colonies in spring relate back to autumn management, particularly the nutrition intake the colonies are exposed to at that time of year.” Take home message: poor nutrition in late summer is tough on the colony, whether varroa is present or not—varroa just makes it worse.
The Effects of Poor Late-Summer Nutrition
Recent research helps us to understand the biological implications of poor nutrition in the hive. Nutritionally-stressed colonies dump excess workers, but by going into survival mode, can often hold the fort until forage again becomes available. But that doesn’t mean that workers reared during periods of nutritional stress are up to par.
The title of a study by Scofield and Mattila [8] summarizes it succinctly: “Honey bee workers that are pollen stressed as larvae become poor foragers and waggle dancers as adults.” Their conclusions are worth noting:
- Mean lifespan was reduced at minimum by 5 days and at most by 18 days for pollen-limited workers compared to workers in control groups, which represents a decrease in mean longevity of 21–56% across trials.
Practical application: shortening the survivability curve of workers leads to an accelerated reduction in the colony population.
- Compared to nestmates who were reared under conditions of pollen abundance, pollen-stressed workers were lighter, they died sooner, and fewer of them were observed foraging. Those who did forage initiated foraging sooner, and foraged for fewer days.
Practical application: reduced foraging ability exacerbates the nutritional stress of the colony.
- Workers reared in pollen-stressed colonies were far more likely to disappear on their first day of foraging (30% vs. 13%).
Practical application: nearly a third of foragers not returning from their first venture outdoors—think about the implications.
- In addition to being less likely to forage, workers reared in pollen-limited colonies were also less likely to waggle dance than control workers and, if they danced, their dances were less precise (although they danced with similar effort).
- These effects suggest a lasting legacy for workers of nutritional stress, one that seriously compromises the foraging and recruitment ability of adults, even when stress is restricted to the larval stage only. It is likely that these behavioral deficits would be exacerbated if chronic stress persisted throughout adulthood at the worker level, and if a greater proportion of workers were undernourished at the colony level.
Varroa adds insult to the injury. van Doormalen [9] found that workers parasitized by varroa during their pupal stage emerged with similar physiology as that of nutritionally-stressed workers, and that they were unable to rebuild their body stores after emergence, even if pollen was abundant. He also suggested that pollen-stressed nurses may sacrifice their own body reserves in order to feed larvae; this would result in shortened longevity of those nurses once they transitioned to foraging duties.
Practical application: it appears that the effect of varroa parasitism of pupae is similar to that of nutritional stress of larvae. So imagine what occurs when a large proportion of the pupae are also parasitized by varroa. Let’s revisit the graph below (Fig. 3).
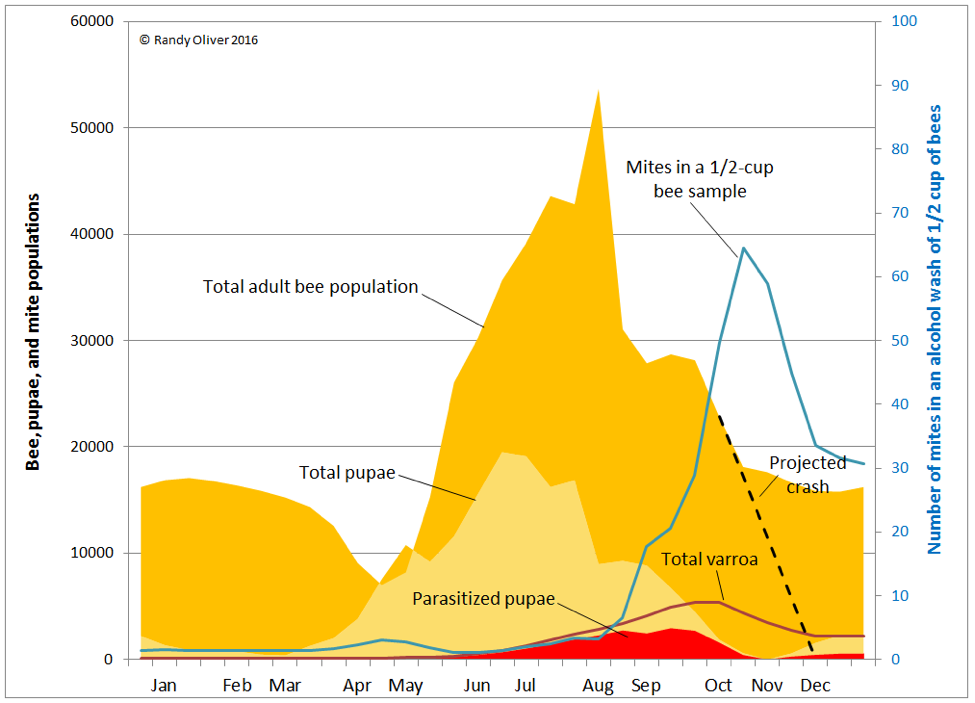
Figure 3. Beekeepers in northern climes (as illustrated here) have it easier, since varroa can only increase when colonies are rearing brood. In contrast, for those beekeepers in southern climes, mite reproduction begins in earnest earlier in the season, and varroa buildup can be far greater than indicated in this graph. Here you can see the amount of varroa hidden in the brood (red), and why alcohol washes suddenly spike in late summer (due to the amount of brood and adult bees rapidly decreasing relative to the number of mites in the hive). What’s important to realize is that even if a beekeeper applies a miticide that blasts all the mites off the adult bees in September [[i]], there is still a huge reservoir of mites in the brood that will soon take their place.
[i] This would include any short-term treatment other than formic acid.
Practical application: it appears that the effect of varroa parasitism of pupae is similar to that of nutritional stress of larvae. So imagine what occurs when a large proportion of the pupae are also parasitized by varroa. Let’s revisit the graph below (Fig. 3).
Figure 3. Beekeepers in northern climes (as illustrated here) have it easier, since varroa can only increase when colonies are rearing brood. In contrast, for those beekeepers in southern climes, mite reproduction begins in earnest earlier in the season, and varroa buildup can be far greater than indicated in this graph. Here you can see the amount of varroa hidden in the brood (red), and why alcohol washes suddenly spike in late summer (due to the amount of brood and adult bees rapidly decreasing relative to the number of mites in the hive). What’s important to realize is that even if a beekeeper applies a miticide that blasts all the mites off the adult bees in September [[i]], there is still a huge reservoir of mites in the brood that will soon take their place.
[i] This would include any short-term treatment other than formic acid.
Even more important than the amount of varroa in the sealed brood is the proportion of the sealed brood parasitized by the mite. Assuming that that the colony begins the season with a very low mite level (100 mites in the example above), by late in the season, a third to more than half of the brood may be infested [11]. Those parasitized workers, should they emerge, will never be the bees that they could have been, thus accelerating the decline of the colony.
Practical application: poor late summer nutrition, coupled with high varroa levels, is a one-two punch that severely weakens the colony, not only in number of bees, but in its resilience against disease—especially that caused by Deformed Wing Virus.
Enter The Viruses
The RNA viruses are endemic in the pollinator population, readily transmitted via contact at flowers, drift of bees between hives, and in the case of DWV, vertically transmitted from queen to egg. Varroa acts as both a vector and initiator of disease from DWV and the paralytic viruses [12]. Without the presence of varroa at a high enough level, these viruses are fairly well tolerated by the colony. But once the infection prevalence in the pupae and adult bees reaches a certain threshold due to vectoring by varroa, the viruses can then go epidemic in the hive.
Practical application: Remember, it’s not varroa that kills colonies, it is the virus epidemics that the mite initiates. Kudos to Dr. Stephen Martin for first elucidating this process [13], and for his vast body of research into understanding bee, varroa, and virus dynamics.
Once the epidemic takes hold, the diseased adults are either driven from the hive [14] or voluntarily commit “altruistic self removal.” Little recruitment of replacement workers can take place, due to the death of the brood from virus infection, inadequate feeding, or chilling. The colony may then suffer through either a heart-rendering dwindling, or the process can snowball into a rapid depopulation leading to what appears to be an overnight collapse [15].
Practical application: the insidious interactive combination of poor nutrition, varroa, and viruses can spell morbidity or death to a colony. And fixing only one problem may not be enough to correct for the others. The poor survivorship of varroa- and nutritionally-stressed bees accelerates the “normal” decline of the colony. Add the altruistic self removal of virus-infected bees and the colony workforce can quickly disappear—leaving behind a hive full of honey, but devoid of bees.
Progression of DWV Collapse
One of the most frustrating things for bee health advisors is beekeeper misdiagnosis of the cause of a colony’s demise when it was clearly due to varroa. In order to help those unfamiliar with the signs of varroa-induced collapse, I’ve included some photos below (Figs. 4-7):
Figure 4. Although this colony is still bursting with bees at the end of the main flow, a DWV epidemic is well underway. Note the discolored and dying propupae and pupae, as well as the workers to the right with deformed wings. Of note is that I often do not see any workers with deformed wings at this stage, but always see dying propupae. Unless something is done to reduce the mite population stat, this colony won’t make it ‘til winter.

Figure 5. It’s only gonna get worse. At this stage, sick fully-developed workers die in their cells. As the undertaker bees remove the carcasses, virions are spread throughout the colony. At this late stage, this colony is essentially one of the “walking dead” although it will still take some time to crash. Even if the beekeeper eliminates all the mites, the virus epidemic has reached the tipping point, and there is little chance for the colony to survive the winter.

Figure 6. In the final stage of collapse, a handful of bees may swarm off with the queen, leaving the remaining workers to drift into other colonies, carrying virus-loaded mites with them. Once the colony can no longer protect itself, robbing bees from other colonies will also carry mites home. Note the telltale mite fecal deposits on the ceilings of the cells, indicative of colony collapse due to excessive varroa.

Figure 7. A close up of the mites’ guanine fecal deposits. To view them in the field, orient the sun at your back [[i]], hold the frame horizontal with the bottom bar toward you, level with your eyes, so that the sunlight illuminates the cells’ “ceilings.” If you see white deposits in the scattered remaining brood cells of a deadout, that’s the signature of collapse due to varroa.
[i] Tip: do not first look to find the sun—it will temporarily blind you. Instead, look for your shadow on the ground and turn so that it is directly in front of you. Sunlight will then come over your shoulder to illuminate the cells.
A surprise finding: A recent study [17] suggests that some forms of DWV are avirulent, and can competitively exclude the virulent strain [18], thus allowing colonies in which the benign form has established a foothold to survive varroa infestation without treatments. This is a very exciting finding, with huge potential for improving colony health.
Practical application: even if the beekeeper manages to wrest control back from varroa, it may take months for a colony to fully regain its health, due to the lingering effects of the virus epidemic. This often comes as a hard lesson to those with almond pollination contracts.
Other Contributing Factors
I’m not forgetting additional potential stressors such as pesticides, miticide-contaminated combs, monoculture pollen, heat stress, etc. Regarding the hot topic of pesticides, when the only (or most attractive) foraging choices in flight range are agricultural crops and surrounding weeds, even sublethal exposures have the potential to exacerbate the problems associated with late summer decline [19].
Practical suggestion: Beekeepers would do well to understand the dilemma that growers (and the EPA) face in attempting to protect our agricultural harvest from pests, while at the same time avoiding unintentional killing of pollinators. For an assessment of the current state of pesticides from an agricultural point of view, I suggest the following article: Insecticides 2016: Out with the Old, In with the New Reality [20].
Any of the above factors (remember “multifactorial”) can make things even worse, or cause problems in their own right. But before pointing any fingers, I’d first make sure that you indeed are providing your hives with good nutrition (natural or supplemental) and truly have varroa under control. I’ll be discussing varroa management in future articles.
Acknowledgements
Thanks to Lloyd Harris for the use of his data set, and to Pete Borst for his generous assistance in library research. And a huge thanks to all the beekeepers who support my research and writing, and give me objective reality reports from all over.
Notes and Citations
[1] One could legitimately still argue that case today. Without varroa, we wouldn’t be pushing $200 for almond rents ($30-$45 in 1997), nor selling nucs for $150 or better ($29 back then). Nevertheless, if given the choice, I’d go back to varroa-free beekeeping in a heartbeat.
[2] In managed apiaries, it is to the benefit of the virus to kill or weaken the colony so that it will be robbed out and transmit the mites carrying that strain to other hives. In a natural situation, this would be nonadaptive once the density of colonies in the environment was reduced. But so long as beekeepers keep restocking deadouts, there is no downside to the virus becoming more virulent. See the paper below for a demonstration of the selection process.
Martin, SJ, et al (2012) Global honey bee viral landscape altered by a parasitic mite. Science 336: 1304-1306. Open access. This is, IMHO, one of the most important studies in explaining what I and others observed in the early years of varroa.
[3] http://www.clemson.edu/extension/beekeepers/factsheets/varroa_mite_control_in_sc.html
[4] Despite commendable efforts by the Baton Rouge Bee Lab researchers. In fact, we’ve actually thwarted the natural selection process that would have already promoted the evolution of mite-resistant bees.
[5] In the early 2000’s there were no registered treatments that still worked. Taktic saved our industry, but it was so cheap and easy to use that many commercial beekeepers never bothered to rotate it with other treatments in order to avoid breeding for resistant mites.
[6] Based upon Lloyd Harris’ data cited previously in this series.
[7] I checked two studies to confirm that pollen income declines in Canadian apiaries at that time of year:
Mattila, HR & GW Otis (2007) Dwindling pollen resources trigger the transition to broodless populations of long-lived honeybees each autumn. Ecological Entomology 32: 496–505.
Nelson, DL, et al (1987) The effect of continuous pollen trapping on sealed brood, honey production and gross income in Northern Alberta. ABJ Aug 1987: 648-650.
[8] Scofield HN, Mattila HR (2015) Honey bee workers that are pollen stressed as larvae become poor foragers and waggle dancers as adults. PLoS ONE 10(4): e0121731.
[9] van Dooremalen C, et al (2013) Interactive effect of reduced pollen availability and Varroa destructor infestation limits growth and protein content of young honey bees. J Insect Physiol. 59: 487–493.
[10] This would include any short-term treatment other than formic acid.
[11] Actual figures are hard to come by—I need to do some brood inspection myself! But the following studies give an idea of the degree of infestation:
Spivak, M & G. Reuter (2001) Varroa destructor infestation in untreated honey bee (Hymenoptera: Apidae) colonies selected for hygienic behavior. J Economic Entomology 94(2):326-331.
Piccirillo, GA & D De Jong (2004) Old honey bee brood combs are more infested by the mite Varroa destructor than are new brood combs. Apidologie 35:359–364.
Calderone NW & LP Kuenen (2001). Effects of Western Honey Bee (Hymenoptera: Apidae) colony, cell type, and larval sex on host acquisition by female Varroa destructor (Acari: Varroidae). J Econ Ent 94: 1022-1030.
[12] Kashmir Bee Virus, Acute Bee Paralysis Virus, and Israeli Acute Paralysis Virus.
[13] Martin, SJ (2001) The role of Varroa and viral pathogens in the collapse of honeybee colonies: a modelling approach. Journal of Applied Ecology 38: 1082 –1093. Open access. This is a necessary read for any serious beekeeper.
[14] Baracchi, D, et al (2012) Evidence for antiseptic behaviour towards sick adult bees in honey bee colonies. Journal of Insect Physiology 58: 1589–1596. The authors found that workers drove asymptomatic DWV-infected bees from the hive. But their data also suggests that that those symptom-free bees also committed altruistic self removal (reviewed in the following paper).
Rueppell, O, et al (2010) Altruistic self removal of health-compromised honey bee workers from their hive. Journal of Evolutionary Biology 23, 1538–1546.
[15] The mechanics are described at https://scientificbeekeeping.com/sick-bees-part-2-a-model-of-colony-collapse/. I developed the model to describe CCD, but the same principles apply to varroa collapse, except that when nosema is not involved, there is little brood remaining at final collapse.
[16] Tip: do not first look to find the sun—it will temporarily blind you. Instead, look for your shadow on the ground and turn so that it is directly in front of you. Sunlight will then come over your shoulder to illuminate the cells.
[17] Mordecai, GJ, et al (2015) Superinfection exclusion in honey bees explains long-term survival of Varroa-infested colonies. The ISME Journal (2015): 1 – 10. Open access.
Mordecai, GJ, et al (2015) Diversity in a honey bee pathogen: first report of athird master variant of the Deformed Wing Virus quasispecies. The ISME Journal (2015): 1 – 10. Open access.
[18] There’s suggestion in other research that the avirulent strain may confer some resistance to nosema. Biology is so complex!
[19] I’m currently testing some remote hive monitors to see whether I can obtain real-time data on the effect of pesticide applications on hives placed in agricultural crops. If successful, I’ll be writing about it.
[20] http://www.agribusinessglobal.com/agrichemicals/insecticides-2016-out-with-the-old-in-with-the-new-reality/


