
Old Bees/ Cold Bees/ No Bees? Part 1

February 24, 2011
The Disease that Disappears. 1
Colony Population Dynamics and Collapse. 3
Dwindling—What a difference a few days make! 5
Foragers have a short warranty. 7
Old Bees/ Cold bees/ No bees?
Part 1
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ in June 2008
“There’s something happening here/ What it is ain’t exactly clear.” This observation by Stephen Stills in 1966 could well describe the state of beekeeping today. Something appears to be amiss in the bee population. There have been an inordinate number of colonies collapsing, sometimes with the workers disappearing “suddenly.” The catch name “CCD” has certainly made the press. Although losses may be due to a variety of problems, the “sudden disappearance” of adult bees may have common denominators.
The Disease that Disappears
“In April 1973 a migratory beekeeper in the Rio Grande Valley of southern Texas…reported the unexplained loss of about 600 of his 3,500 colonies…. The colonies had reared brood earlier in the year when natural nectar and pollen were available. Then when the weather turned unseasonably cold…and rainy for about two weeks, the population of adult worker bees had declined rapidly to a “handful” of workers and a queen; there were only a few dozen dead adults on the bottom board and in front of each colony. Despite more than 30 years of successful beekeeping, this beekeeper was unable to control the declining populations by feeding sucrose syrup and pollen supplement for several weeks.” (Wilson and Menapace 1979).
Dr Wilson reviewed the literature on what was then called “Disappearing Disease,” and found unexplained losses of bees dating back at least to the 1800’s, and noted that A.I. Root had used the term “spring dwindling” to describe losses that occurred during cold weather from February to June. No one was able to explain the phenomenon.
A decade after Wilson’s report, Drs. Lulincevic, Rothenbuler, and Rinderer (1983) again described Disappearing Disease, and again the best minds in the business were unable to determine the culprit. And again, the “disease” seemed to disappear.
Last fall, Brett Adee helplessly watched tens of thousands of his colonies collapse in California, commencing with the first cold spell. I spoke with Brett at the time. I’m still haunted by the frustration in his voice as he described watching his bees die colony after colony. The Adees are no amateur beekeepers, yet found themselves helpless to stop the losses.
Again, some of the best scientific minds are currently working on solving the current collapses that we are experiencing. Dr Gard Otis’ article in the December issue of this Journal detailed some of the issues involved. There has been plenty of “armchair quarterbacking” by beekeepers and bloggers, often without supporting data. I personally do not presume to be in a better position to finger a culprit, especially after having spent time speaking with most of the major CCD researchers. However, I will attempt to clarify several of the “generic” factors involved in colony dwindling, and to explain the possible contributions by the various suspects. As always, the point of my articles is to try to summarize and explain the biological research, so that beekeepers can then think for themselves from an informed standpoint.
I will begin this series of articles with a discussion of factors that can cause dwindling or collapse of a colony. In subsequent articles, I plan to discuss “the usual suspects,” with a special focus on the honey bee viruses, an introduction to the bee immune system and its implications in bee husbandry, and finally a look at bee genetics, and an examination of the strategies that we have used for bee breeding in this country. I am well aware that this is an ambitious undertaking, but I feel that the bee industry is at a point where we need to reevaluate the basic science behind everything we do or don’t do.
The Disappearing Act
Unlike colonies dwindling from bee brood diseases (such as AFB, EFB, and chalkbrood), which are generally easy to spot, unexpected “sudden” colony collapses are due to the loss of adult bees. Note that I use the term “loss,” rather than “disappearance,” of adult bees. There have been hypotheses that cell phones or insecticides are causing the bees to “lose their way” home. So let’s look at the plausibility of “disoriented bees.”
The Adees lost some 25,000 colonies (nearly 40% of their operation) in California prior to almond bloom. Assuming a conservative average of 6-frame strength, and 1800 bees per frame, that is 270,000,000 bees, or nearly 40 tons of bees! The Adee bees were in large holding yards—groups of white boxes that stood out starkly in the gently rolling grass-covered hills. To think that 40 tons of bees couldn’t find or smell their ways home to the only landmarks around is rather implausible!
What more likely happened was that the adult bees flew out, and were unable to return for some reason. Clearly we need to focus our attention upon what factors might prevent bees from returning.
Update 4/22/2011 Since I wrote this article, it’s become clear that sick bees may intentionally abandon the hive, in a process termed “altruistic self removal.” We’ve also learned that Nosema ceranae infection may result in the loss of bee blood storage sugar (and thus a bee’s ability to warm the flight muscles).
Colony Population Dynamics and Collapse
Normal colony growth is to build up in spring, then swarm, then build up again through midsummer. The queen’s egglaying peaks generally in July, and the colony population starts to drop down again in late summer and fall (see Figure 1).
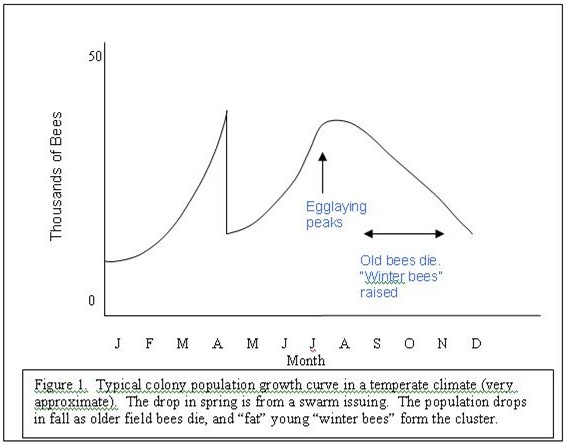
The loss of large numbers of older field bees in fall is normal, and up to half the population may be lost in preparation for winter. This is a healthy, normal process, since it likely removes disease-carrying older bees from the wintering cluster, and prevents piles of corpses from overwhelming the colony during winter. The foragers wouldn’t live long anyway, since they had already given up the bee “fountain of youth”—vitellogenin—when they switched jobs from being house bees to working as foragers (Seehus 2006).
As long as bees are loaded with vitellogenin, they can live for a long time—years as a queen, or for as much as ten months as “winter bees” in the far north! However, when vitellogenin-rich house bees switch to foraging, they apparently feed their vitellogenin reserves back to the others, so as not to take unnecessary protein from the colony when they take on the risky job of foraging. Unfortunately, without vitellogenin reserves, they sacrifice much of their immune response, and free-radical scavenging ability, and thus begin aging (Amdam 2003).
The old worn-out foragers wouldn’t be of much use to the colony during winter, and could actually be a detriment if they were loaded with viruses or nosema. So they simply work themselves until they wear out in the field, or possibly fly off to perform “altruistic suicide.” The remaining winter cluster then consists of healthy, “fat” winter bees, full of vitellogenin, which helps to protect them from aging due to oxidation reactions.
The colony still has more tricks up its collective sleeve! By clustering over any remaining brood, and allowing the outer shell of the cluster to drop to 41ºF, the bees can conserve their energy and food stores until needed for broodrearing after the winter solstice. The cluster core has an incredible ability to generate heat—it can produce a metabolic rate equal to that of a shrew (Kronenberg & Heller 1982)! However, the bees make every effort instead to conserve energy, and trap the metabolic heat from the core by tightly packing their bodies into an efficient insulation.
Even more remarkable, van Nerum & Buelens (1997) describe how the colony conserves energy and water by reducing ventilation with the tight winter cluster. The bees allow carbon dioxide levels to rise to as high as 6% (5% is dangerous to humans) inside the cluster. At the same time they create a condition of hypoxia (low oxygen), reducing the oxygen level from the 21% in the atmosphere to about 15%, which induces them to enter into an “ultra low metabolic rate”—similar to the diapause that other insects use to survive the winter. In this state, the colony uses minimum energy, and traps critical metabolic water within the cluster (the center experiences low relative humidity, and the cooler shell traps escaping moisture). (No mention was made of the effect that such an extraordinarily high CO2 level would have in increasing the acidity of the haemolymph—this may be an effect deserving a bit of research).
The authors may have overlooked yet another benefit to hypoxia—to avoid oxidative aging (Fridovich 1977). The bee respiratory system is extremely effective at diffusing oxygen into the tissues—some 200,000 times faster than our human delivery system that uses blood (Burmester 2005). He states that “oxygen is a double-edged sword: although required to fuel energy production, it is also a potent source of toxic compounds known as reactive oxygen species (ROS), which can damage proteins, DNA and lipids. In recent years, ROS have been recognized as a major threat to cell survival, and toxic ROS effects are suggested to underlie ageing and cell death. Therefore, it is advantageous to keep cellular oxygen levels just high enough for efficient mitochondrial respiration, and as low as possible to minimize oxidative damage.”
“The insects’ tracheal system is well designed for efficient oxygen supply during periods of high activity [such as flight], when oxygen never accumulates to critical concentrations in the cell because it is rapidly converted into water by the respiratory chain. However, in periods when respiration falls, such as in [a winter cluster], oxygen consumption is too low to prevent it building up to harmful levels.”
Furthermore, Hetz (2005) states that “increases in the level of O2 in the atmosphere that…insects are breathing lead to enhanced rates of oxidative damage and reduced longevity. Oxygen has been shown to be a toxic molecule that is needed for oxidative metabolism but must be supplied in carefully controlled amounts and concentrations.” And this is just what the winter bee cluster appears to do!
So winter bees have two tricks to use to keep from aging during a cold winter: keeping oxygen levels low, and using vitellogenin to scavenge harmful free radicals. Van Nerum & Buelens suggest that the low-oxygen principle explains why very small or very large colonies may have trouble surviving the winter. The also point out that any wintertime disturbance of the cluster during cold conditions can be detrimental to a surprising degree. This certainly makes me curious as to the effect of winter trucking of bees to almonds, and may explain why they occasionally collapse shortly after they are unloaded.
Dwindling—What a difference a few days make!
OK, so much for normal colony growth and wintering. Let’s talk about dwindling populations. Total colony population is a simple function of bee birth rate minus bee death rate. If the queen lays 1500 eggs a day, and most emerge as adults 21 days later, the colony will experience a “recruitment” of 1500 bees per day. This potential growth is offset by the rate of death of bees as they become foragers and die (or, more specifically, fail to return). If more bees emerge each day than fail to return, the colony will grow. If they are lost faster than they emerge, then the colony size will shrink.
In the collapses that are being reported, the problem does not appear to be a low birth rate, since there is (or was) ample (apparently) healthy brood present. So we’d better look instead at death rate.
Since most bees spend about three weeks as house bees during the spring and summer (Seeley 1995), the critical variable is how many days they survive as foragers (or more properly, the average number of days until they fail to return). Refer to the two graphs below. I used Dr. Gloria DeGrandi Hoffman’s improved VarroaPop modeling program (available for free download). In the first graph (Figure 2), I set forager lifespan at 16 days; in Figure 3 at 4 days.
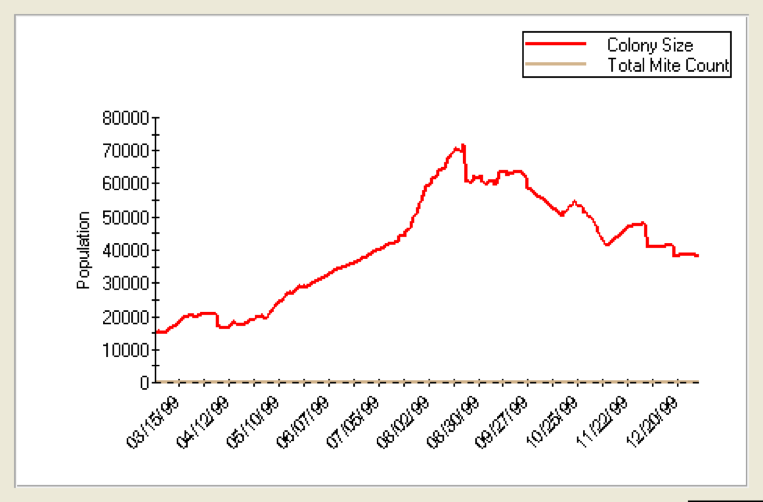 Figure 2. VarroaPop run to model colony growth in the Southwest, starting with 8 frames of bees with brood, a moderate queen, no mites, and forager longevity set at 16 days. The population reaches 70,000 bees, and runs about 8 frames of brood (not shown) during summer. This strong colony winters with 20 frames of bees.
Figure 2. VarroaPop run to model colony growth in the Southwest, starting with 8 frames of bees with brood, a moderate queen, no mites, and forager longevity set at 16 days. The population reaches 70,000 bees, and runs about 8 frames of brood (not shown) during summer. This strong colony winters with 20 frames of bees.
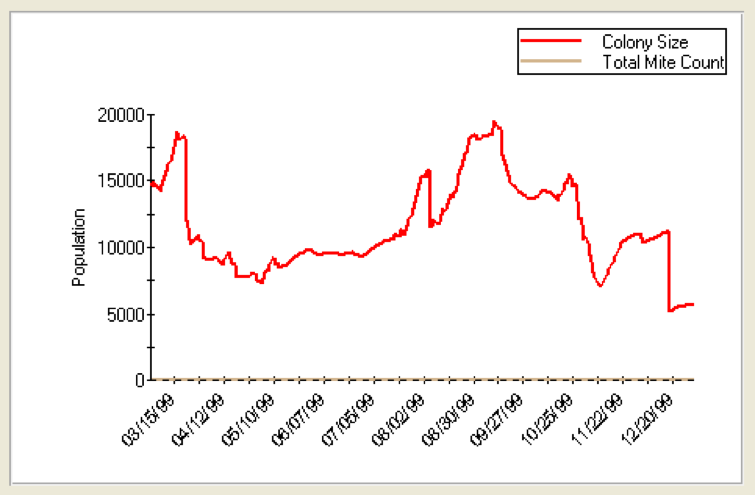
Figure 3. VarroaPop run using same parameters, except forager longevity reduced to 4 days. This colony never reaches 20,000 bees! (unfortunately the program rescales the vertical axis, so the graphs are a bit hard to compare visually). This colony experiences a “spring dwindling” as workers begin foraging (and thus aging). However, if you were to inspect the colony on any day, it would look healthy, but lagging, since it would have about 4 frames of brood, plus a good population of “house bees”—but it would only build up to about 11 frames of bees. This colony then dwindles to about 4½ frames in fall, and then drops to about 3 frames of bees during winter. (If the simulation is continued into the next season, the colony never recovers.)
As you can see, a shortened forager average lifespan has a striking effect upon colony population. If foragers live for 16 days, the colony population builds robustly as long as there is pollen available. However, if foragers only live for 4 days, the colony is in trouble. Although the colony would appear to be healthy upon inspection, in actuality it would be struggling to stay alive! All that the beekeeper would notice would be a “lagging” colony that might suffer from both spring and fall dwindling, not build up well despite having frames of healthy of brood, and collapse during the winter for no apparent reason.
Premature loss of foragers has a profound effect on a colony beyond population size. Fewer foragers means less food in the hive, and poorer nutrition for the brood and nurse bees. Young bees compensate by initiating foraging earlier in life, which then depletes the number of nurses available to feed the queen and brood.
It thus appears that the key factor that causes the dwindling of apparently healthy colonies is how many days the average forager lives until it fails to return. That being so, we should then look at some of the factors that can decrease forager survivability. Many are of no great mystery. However, the beekeeper may not notice them at play!
Foragers have a short warranty
Just what causes a bee to become “old”? Adult bees move through four main stages of behavior, starting with jobs within the broodnest, then two phases working toward the periphery of the cluster, and finally working as foragers (Seeley 1995). This progress is determined by genetics, physiological development, nutrition and hormone titers (Nelson 2007), and colony needs. The aging of bees is in interesting topic, as it is more physiological than chronological. House bees are physiologically prepared to forage by about the age of eight days (Whitfield 2006), but generally wait until they are much older to leave the hive (about 6 weeks in spring, or 3 weeks in summer (Neukirch 1982).
As I mentioned earlier, the aging of bees doesn’t begin until they become foragers. So let’s look at the implications of this. A general rule in biology is that animals that “expect” to live a long time (think turtles) are rewarded by investing precious energy and resources into the maintenance and repair of their body tissues, whereas animals that are unlikely to live very long due to predation or loss of habitat (think mice) forego maintenance, and instead invest their energy into reproduction. They thus tend to “wear out” and die at an early age, even under the best of circumstances.
Bees follow a dual pattern: nurse bees or “winter” bees (also handily called “diutinus” bees) have few external pressures that would result in death, so they invest in maintenance of their bodies by keeping their vitellogenin and glycogen levels high, and their immune systems strong. This gives this them greater resistance to stressors such as starvation, thermal, and oxidative stress (Remolina 2007). “Diutinus bees stay in the nest and survive unfavorable periods such as winter…. They differentiate into nurse bees and foragers when conditions improve and brood rearing commences” (Seehuus 2007).
However, as soon as they switch to foraging, they now have a finite life expectancy due to a single wear item—their wings (Figure 4). An insect is unable to repair its tattered wing tips, just as we can’t repair a worn tire on our cars. As an analogy, imagine that you were on a desert island, and owned an automobile with only one non replaceable set of tires. There would be no reason for you to maintain the engine beyond what it would take to make it last for the expected life of the tires. And there is no reason for a forager to maintain its flight motor beyond the expected life of its wings. Nature has apparently figured out the math, and programmed the forager’s flight muscles to be able to sustain about 150-500 cumulative flight miles (Neukirch 1982).
Several mechanisms are involved in the actual “wearing out” of the forager’s motor (this is a well-debated topic (Speakman 2005), which I do not presume to do justice to). The flight muscles do not maintain their mitochondria (Tribe 1972), lose their ability to recharge glycogen (Corona 2005), and suffer from oxidative stress. Plus, as the wings get shredded, the poor bees have to beat their wings faster and faster just to fly (Higginson 2004).

Figure 4. An aged forager bee on fennel blossoms. Note the worn, ragged wings. The wings are a non repairable part of the bee, and thus limit functional life as a forager. Photo © the author.
Couple this hard work with cumulative exposure to pesticides (more on this later), pollution, and exuberant use of miticides, and forager lifespan can be greatly truncated.
In cold weather, this problem is exacerbated, since foragers must shiver to keep their flight muscles warm (more on this later), and thus they may “wear out” even more quickly. Foraging in cool weather would be expected to accelerate the “aging” of bees.
Foragers are thus an expendable element of a colony. There are no “golden years” for old bees–they use all their limited resources to seek out and gather pollen, nectar, water, or propolis, and simply work themselves until they drop. The harder they have to work, the sooner they wear out. When apiaries contain more colonies than the immediate pasture will support, or if the water source is distant, the foragers will wear out in fewer days since more travel is involved in picking up the groceries.
The take home message on bee aging is that it appears that a bee has a finite number of wingbeats to spend. As long as she stays inside and keeps her vitellogenin titer up, she can live a long time. But once she begins flying, she’s got a maximum 500-mile warranty. There are various figures cited, but that works out to be about 8-12 flight days for a healthy, well-nourished forager, which brings us to…[click here for Old Bees Part 2 ]
References
Allsopp, M (2008) Tracheal Mites.(Broken Link!) http://www.arc.agric.za/home.asp?PID=1&ToolID=63&ItemID=3081
Amdam, G. V., K. Norberg, A. Hagen, and S. W. Omholt (2003) Social exploitation of vitellogenin. Proc. Natl. Acad. Sci. U.S.A. 100: 1799Ð1802.
Bailey, L & EFW Fernando (1972) Effects of sacbrood virus on adult honey-bees. Ann. Appl. Biol. 72:27-35.
Carreck, N (2008) CCD – A View From Across The Pond. Bee Culture Jan 2008
Berényi, O, T Bakonyi, I Derakhshifar, H Köglberger, N Nowotny (2006) Occurrence
of six honeybee virus in diseased Austrian apiaries. Appl Environ Microbiol 72, 2414-2420.
Bromenshenk, J (2008) pers comm.
Burmester, T. 2005. A welcome shortage of breath. Nature. 433: 471-472
Corona, M, KA. Hughes, DB Weaverd and GE Robinson (2005) Gene expression patterns associated with queen honey bee longevity. Mechanisms of Ageing and Development 126(11): 1230-1238
Cox-Foster, D, et al. (2007) A metagenomic survey of microbes in honey bee colony collapse disorder. Science 318(5848): 283-287.
Cox-Foster, D, et al (2007) Supporting Online Material for A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. www.sciencemag.org/cgi/content/full/1146498/DC1
Dall, D. J. (1985). Inapparent infection of honey bee pupae by Kashmir and sacbrood bee viruses in Australia. Ann Appl Biol 106, 461–468.
Fisher, FM Jr and RC Sanborn (1964) Nosema as a source of juvenile hormone in parasitized insects. Biol Bull 126: 235-252.
Fluri, P, M. Lüscher, H. Wille and L. Gerig (1982) Changes in weight of the pharyngeal gland and haemolymph titres of juvenile hormone, protein and vitellogenin in worker honey bees Journal of Insect Physiology Volume 28, Issue 1, Pages 61-68
Fridovich, I. 1977. Oxygen is toxic! Bioscience 27: 462-466.
Goodwin, R.M., Taylor, M.A. (2003) Diagnosis of Common Honey Bee Brood Diseases and Parasitic Mite Syndrome. (Broken Link!) http://www.hortresearch.co.nz/files/science/biosecurity/227525-Bee-Pamphletpths-small.pdf
Heinrich, B (1996) The Thermal Warriors. Harvard University Press
Heinrich, B (2004) Bumblebee Economics. Harvard University Press
Hetz, S.K. and T.J. Bradley. 2005. Insects breathe discontinuously to avoid oxygen toxicity. Nature 433: 516-519.
Higginson, AD and FS Gilbert (2004) Paying for nectar with wingbeats: a new model of honeybee foraging Proc Biol Sci. 271(1557):2595-603.
Kleinhenz, M, B Bujok, S Fuchs and J Tautz (2003) Hot bees in empty broodnest cells: heating from within. The Journal of Experimental Biology 206, 4217-4231.
Köglberger, H, I Derakhshifar, J Kolodziejek, H Homola, and N Nowotny (2006) Prevalence of six honeybee viruses in beehives collected at different Austrian locations during different seasons, and correlation with non-viral disease. Proceedings of the Second European Conference of Apidology EurBee
Kralj J, S. Fuchs, J. Tautz (2006) Disease removal by altered flight behavior of forager honey bees (Apis mellifera) infested with Nosema apis. Proceedings of the Second European Conference of Apidology EurBee
Kronenberg, F & HC Heller (1982) Colonial thermoregulation in honey bees (Apis mellifera) -. J Comp Physiol 148:65-76
Kulincevic, JM, WC Rothenbuhler, TE Rinderer (1984) Disappearing disease: III. A comparison of seven different stocks of the honey bee (Apis mellifera). Research bulletin 1160, Ohio State University
Manning, R (2008) The Effect of High and Low Fat Pollens on Honeybee Longevity. RIRDC Publication No 08/031.
Matilla, RM and GW Otis (2006) The effects of pollen availability during larval development on the behaviour and physiology of spring-reared honey bee workers. Apidologie 37:533–546
Schmickl, K & K Crailsheim (2004) Inner nest homeostasis in a changing environment with special emphasis on honey bee brood nursing and pollen supply. Apidologie 35: 249–263
Seehuus, SC, et al. (2006) Reproductive protein protects functionally sterile honey bee workers from oxidative stress. PNAS 103(4): 962-967.
Shen, M L Cui, N Ostiguy and D Cox-Foster (2005) Intricate transmission routes and interactions between picorna-like viruses (Kashmir bee virus and sacbrood virus) with the honeybee host and the parasitic varroa mite. Journal of General Virology 86: 2281–2289
Nelson, CM, KE Ihle, MK Fondrk, RE Page Jr., GV Amdam (2007) The Gene vitellogenin Has Multiple Coordinating Effects on Social Organization. Public Library of Science http://biology.plosjournals.org/perlserv/?request=get-document&doi=10.1371/journal.pbio.0050062
Neukirch, A (1982) Dependence of the life span of the honeybee (Apis mellifica) upon flight performance and energy consumption. Journal of Comparative Physiology 146(1): 35-40.
Nielsen, SL, M Nicolaisen, P Kryger (2008) Incidence of acute bee paralysis virus, black queen cell virus, chronic bee paralysis virus, deformed wing virus, Kashmir bee virus and sacbrood virus in honey bees (Apis mellifera) in Denmark. Apidologie 39 DOI: 10.1051/apido:2008007
Remolina SC, Hafez DM, Robinson GE, Hughes KA (2007) Senescence in the worker honey bee Apis mellifera. J Insect Physiol. 53(10):1027-33.
Seehuus S, Norberg K, Krekling T, Fondrk K, Amdam GV. (2007) Immunogold localization of vitellogenin in the ovaries, hypopharyngeal glands and head fat bodies of honeybee workers, Apis mellifera. Journal of Insect Science 7:52, available online: insectscience.org/7.52
Seeley, T (1995) The Wisdom of the Hive. Harvard University Press.
Speakman, JR (2005) Body size, energy metabolism and lifespan. Journal of Experimental Biology 208, 1717-1730
Stills, S. (1966) For what it’s worth. Recorded by Buffalo Springfield
Stoffolano, J (nd) Respiratory or ventilatory system http://www.faculty.ucr.edu/~insects/pages/teachingresources/stoffolano/14.pdf
Southwick, E (1992) The physiology and social physiology of the honey bee. In The Hive and the Honey Bee. Dadant and Sons
Tentcheva, D, L Gauthier, N Zappulla, B Dainat, F Cousserans, M E Colin, and M Bergoin (2004) Prevalence and Seasonal Variations of Six Bee Viruses in Apis mellifera L. and Varroa destructor Mite Populations in France. Applied and Environmental Microbiology, December 2004, p. 7185-7191, Vol. 70, No. 12
Tribe, MA and DE Ashhurst (1972) Biochemical and structural variations in the flight muscle mitochondria of aging blowflies, Calliphora erythrocephala. J. Cell Sci. 10: 443-469
Van Engelsdorp, D, D Cox Foster, M Frazier , N Ostiguy, J Hayes (2006) “Fall-Dwindle Disease”: Investigations into the causes of sudden and alarming colony losses experienced by beekeepers in the fall of 2006.


