First published in: American Bee Journal, April 2014
CONTENTS
Setting The Stage: The Origins Of The Players
Early Changes in The Honey Bee Niche Due To Humans
The Human Deforestation Of Europe
Adaptation And Change In Business
The Creation Of A Niche For Bee-keeping
The Domestication Of The Honey Bee
The Price Of Domestication
Acknowledgements
Citations and Footnotes
What’s Happening to the Bees?
Part 3
Originally published in ABJ April 2014
Randy Oliver
ScientificBeekeeping.com
In the last installment of this article I explored the limiting factors of the honey bee realized niche prior to the influence of humans. So let’s now look at how the populating of Europe by modern humans affected the honey bee.
Setting The Stage: The Origins Of The Players
Let’s first set the stage. The time frame of interest runs from about 10,000 years ago through about 400 years ago. This period spans the time from the last “ice age” (technically, the glacial period; during which it was too cold for bees to inhabit the area), covers the invasion and colonization of the warming continent by both bees and modern humans, and ends with when humans started transporting the honey bee across oceans.
Apis mellifera
The honey bee evolved long prior to the time frame of interest. Recent research [1] suggests that the genus Apis originally developed in Europe, and then spread into Asia (where it evolved into several different species), into North America (where it later went extinct), and into Africa (via Spain/Morocco). It was in Africa that the species Apis mellifera evolved, and then later migrated back into Europe and the Middle East, branching in various ecological habitats (realized niches) into locally-adapted subspecies.
Then came a prolonged Ice Age, during which there were periods of cold and dry glaciation interspersed by periods of warming (as we are currently enjoying). During the cold periods, ice covered northern Europe, and honey bees were forced to follow suitable habitat southward, retreating into warmer “refugia.” During the warm (and wetter) periods, ice retreated and Europe could temporarily revegetate, allowing honey bees to expand their ranges back again northward. The current races of bees in Europe recolonized the region from such refugia as the climate warmed about 10,000 years ago [2]. I’ve shown the distribution of the named races (subspecies) of Apis mellifera in the map below (Fig. 1). The satellite image shows the differences in climate and vegetation in the various regions. Of note is that the first “keeping” of bees appears to have begun in the Fertile Crescent of the Middle East with A. m. jemenitica and syriaca.
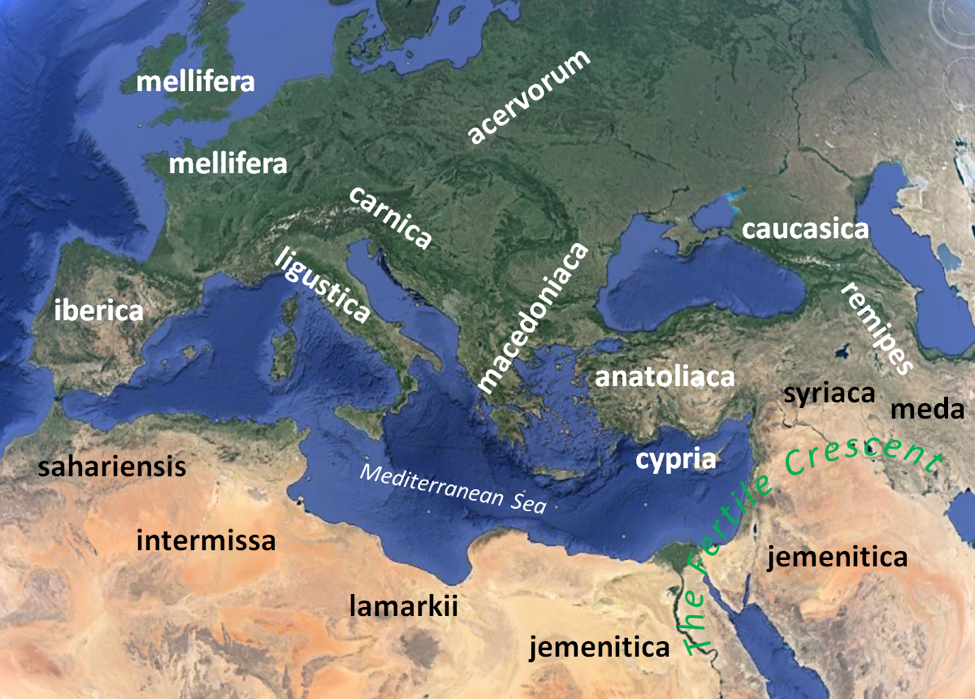
Figure 1. Subspecies of Apis mellifera in Europe and the Mediterranean region. Our domesticated stocks in the U.S. primarily derive from the temperate-adapted ligustica (Italian), carnica (Carniolan), and perhaps caucasica. The feral population of bees in the U.S. also contains the mellifera (German dark) lineage [[i]]. Map by the author; satellite image from Google Earth; subspecies distribution from various sources [[ii]].
[i] Delaney, D (2008) Genetic characterization of U.S. honey bee populations. PhD Thesis http://www.dissertations.wsu.edu/Dissertations/Summer2008/d_delaney_070108.pdf
[ii] The classification of these races is rather arbitrary, and under debate by taxonomists, but gives a general idea. There are also discrete breeding populations within each of the subspecies. See:
Radloff, SE, and HR Hepburn (1998) The matter of sampling distance and confidence levels in the subspecific classification of honeybees, Apis mellifera L. Apidologie 29: 491-501. Open access.
Al-Ghamdi, AA, et al (2013) Geographical distribution and population variation of Apis mellifera jemenitica Ruttner. Journal of Apicultural Research 52(3): 124-133. Open access.
Practical application: each subspecies of honey bee is adapted to a specific habitat and climate. The “best” bee for any region is that which has already undergone countless generations of adaptation. Although the Italian bee is very popular among many beekeepers, it is certainly not the best adapted bee for non Mediterranean [5] conditions.
Homo sapiens
At the end of the last glacial period, modern humans also moved up into Europe from Africa, displacing the cold-adapted Neanderthals (as did the bees, modern humans also evolved into different races in the region). These hunter-gatherer human populations were at first not dense enough to exert an appreciable impact on the honey bee. This changed with the advent of pastoralism and agriculture—initially slash and burn, then later improved by the invention of the plow. The adoption of agricultural practices facilitated the niche of Homo sapiens—by greatly increasing the carrying capacity of the habitat (mainly by the farming of grain)–thus allowing the human population to begin its exponential growth (to be later limited by epidemics of infectious diseases).
Practical application: as we shall see, the factors of migration, locally-adapted races, displacement of existing populations, the farming of grain, and epidemics of infectious diseases will greatly affect the honey bee over the ensuing years.
Early Changes In The Honey Bee Niche Due To Humans
O.K., now that I’ve set the stage, let’s take a look at how the early expansion of humanity into the native range of the honey bee affected the limiting factors of the bee’s niche (I will cover more recent impacts later). Allow me to address each of the limiting factors that I’ve previously covered, in turn.
Limiting factor: The weather
Weather is the day-to-day expression of climate. For hundreds of thousands of years, the climate of Europe oscillated between periods of cold/dry and warmer/wetter, which of course greatly affected the local weather. The fundamental niche of the honey bee is limited by cold and prolonged winters, by extreme summer heat, and by lack of water and nectar-producing plants (especially forbs—herbaceous flowering plants other than grasses). For thousands of years at a time, parts of Europe simply did not provide conditions that met the requirements of the bees’ fundamental niche, causing the extirpation of local populations or entire species. Keep in mind that any species has “edges” to its range, past which the species is stressed, or cannot successfully live. Slight changes in weather at the edges can temporarily make that habitat unsuitable for bee survival.
Practical application: for example, cold, wet summers in England may not allow bees to store enough honey to make it through the winter. Ditto for drought-prone California. And an unusually severe northern winter will challenge colonies of Italian bees.
We humans have little ability, other than by fervent rain dancing, of changing tomorrow’s weather; however, we do have the ability to change the climate on a local basis, and likely even at the global scale. Climate then may affect the weather.
The burgeoning human population in Europe and the Mediterranean started grazing herds of domesticated mammals and cutting down the forests [6]. This loss of the shading forest cover likely resulted in the warming of central Europe, and the desertification of the Mediterranean region [7]. Such deforestation likely favored bees in central Europe (due to creating better conditions for forbs), but created drier (and less favorable) microclimates in the Mediterranean.
The Human Deforestation Of Europe
Let’s look at the vegetation of Europe at the beginning of this period of time (Fig. 2):
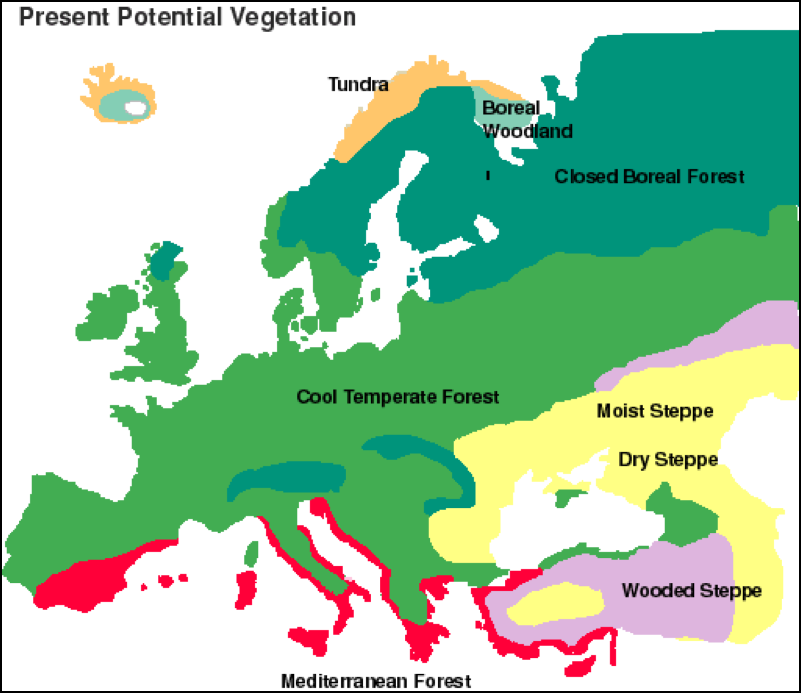
Figure 2. The vegetation of Europe around 4,500 years ago, just before the main agricultural and deforestation phase by humanity in the region [[i]].
[i] Map from Adams, op. cit.
Note the extensive forest cover in the natural range of the honey bee in Europe and the Mediterranean at this time in geological history. Not shown is the range of the honey bee in the moister areas of the Middle East and northern Africa, which were also densely forested. Adams [9], by reviewing data on fossils of pollen, tracked the destruction of these forests by humans over the course of a few thousand years (accelerated about 3000 years ago by the invention of the iron axe and saw). The deforestation of the habitat brought about major changes in two limiting factors of the niche of the honey bee—the abundance of forage, and the availability of nest cavities.
Limiting factor: Carrying capacity of the habitat
Although some trees provide pollen in the spring, and a few, nectar, dense forest is not prime honey bee habitat, since the tree canopy shades out flowering low-growing forbs and shrubs. It was only in natural meadows and openings of such forest that there would have been suitable forage for bees over most of the season. However, such ancient trees would have furnished abundant nest cavities.
When humans invaded those areas, they practiced slash and burn agriculture, clearing the forests for pasture or crops, or cutting trees for structures, monuments, shipbuilding, or charcoal. These forests were largely devastated by the end of the Roman Empire. Although we abhor such devastation of virgin forests today, it was likely of benefit to the honey bee, as it allowed sunlight to hit the ground, favoring the growth of bee-friendly forage plants in the pastures and cropland (remember, herbicides had not yet been invented) (Fig. 3).

Figure 3. When I’m asked to give presentations to local groups interested in gardening for the benefit of pollinators, I like to open with this slide to illustrate a point–that by cutting down pine trees (which are of no value to honey bees), one allows sunlight to hit the understory of flowering plants. Since we have suppressed natural wildfires in California, formerly open land is being reclaimed by dense pine and oak forests. Such change has been detailed for my county at [[i]].
[i] Walker, PA, et al (2003) Landscape changes in Nevada County reflect social and ecological transitions. California Agriculture 57(4): 115-121.v
Thus, by clearing the forests (by approximately 75% in central Europe), humans improved (facilitated) one aspect of the realized niche of the honey bee, since such clearing favored the growth of a greater abundance of forage plants. But there was a flip side to this.
Limiting factor: Predation
Humans (by virtue of possessing a sweet tooth, climbing ability, and wood-cutting tools) are a formidable predator of the bee. This may well be one of the reasons that the Savannah Bee (Apis mellifera scutellata) so fiercely protect their nests (their long exposure to human predation would have selected for those colonies which were able to successfully deter human honey hunters).
Hunter-gatherers do not waste energy on hunting prey that does not give a positive return on investment. It takes a considerable investment in energy, pain, and risk of life and limb to harvest the combs from a small-entranced cavity high in a hollow tree (this may be one reason that European bees prefer to nest high in trees [11]. It was only once humans had their bellies full of grain that they had the luxury of satisfying their sweet tooth by making serious efforts to attack well-defended colonies high above ground.
As the human population became more dense, the pressure of predation on the honey bee would have increased greatly, favoring the survival of bees that possessed three traits—cryptic and inaccessible nesting, vigorous defense of that nest, and frequent swarming so that colony reproduction was greater than the loss due to human predation.
Limiting factor: Nesting cavities
The clearing of ancient forests affected another parameter of the honey bee niche. The falling of each hollow bee tree eliminated one available nest cavity. As hollow trees became rarer and rarer (and tended to remain rare in regrown managed forests), there would have been fewer and fewer places for honey bees to nest. The few remaining “bee trees” would have been targeted by honey hunters, who, with the use of steel axes and saws found it easier to simply fall a tree than to climb it. Each of these destructive predations by humans eliminated yet another increasingly rare nest cavity.
So by this time point in history, two major factors of the realized niche of honey bee had been altered by humans—there would have been more herbaceous and shrub forage available, but fewer nest sites. And such change created opportunity for humans to adapt from being honey hunters to honey farmers.
Adaptation And Change In Business
On two occasions in recent years, speakers [12] have suggested at conferences that those of us in the bee business read the motivational booklet Who Moved My Cheese? [13]. I recently did so. It’s a cute little parable that can be read in minutes, but summarizes important lessons in recognizing business opportunities and adapting to changes in business niches. Two of these lessons are to:
Anticipate and Monitor Change, and then
Adapt To Change Quickly
Like it or not, things change. Niches, whether ecological or in business, change continually. Those who adapt may enjoy success; those who don’t, go extinct. Both the honey bee and their keepers have learned to exploit various realized niches, and those niches change over the years. As I mentioned before, both bees and humans are highly adaptive species. Honey bees adapt by the process of genetic (and epigenetic) trial and error that we call evolution. Human beekeepers, generally blessed with larger brains, have the capacity to recognize upcoming changes in their niche, and the associated pitfalls and business opportunities.
However, human nature is such that many will waste their time lamenting about how difficult or impossible change is, rather than quickly adapting. On the other hand, those who are innovative and cognizant of business opportunities consistently make money.
Let me state emphatically that I do not consider myself to be any sort of great beekeeper or business guru. But what I have noticed over the years are inherent differences in the business attitudes of those beekeepers who always seem to be complaining, compared to those who are able to afford shiny new trucks. Throughout this article I will return to adaptations in the business of keeping bees made by successful beekeepers. So let’s return to the change in the niche of traditional honey hunters in the homeland of the European honey bee.
The Creation Of A Niche For Bee-keeping
Honey hunters would have put themselves out of business once they cut down all the hollow trees. That situation created a novel business opportunity, since there would now exist an insufficient number of natural nest sites for the number of colonies that could be supported by the local forage. All that a human entrepreneur needed to do would be to facilitate the bees’ realized ecological niche by supplying them with what had now become the major limiting factor—the lack of suitable nest cavities. And voilà, as the business niche of honey hunting dwindled, those skilled at plundering bee trees could adapt to become…bee-keepers! This supplying of nest cavities would have been especially successful in the Fertile Crescent once it lost its forests, and would also have allowed bee-keepers to expand the honey bee’s range into arid areas naturally lacking trees or rock cavities.

This beekeeper in Yemen supplies his bees with nest cavities in a landscape lacking such naturally. Honey from Yemen fetches a high price—over $100 per pound by mail order [[i]]. The growing popularity of beekeeping in Yemen today suggests that the beekeepers there may soon reach the carrying capacity of the land [[ii]]. Photo by Gillian Duncan [[iii]].
[i] http://www.balqees.com/shop/yemeni/
[ii] http://www.yementimes.com/en/1633/business/1736/Liquid-pot-of-gold-Yemen’s-honey-trade.htm
[iii] http://www.thenational.ae/business/industry-insights/the-life/sticky-patch-for-yemen-honey-exports
The honey bee, when kept as livestock, exhibited a trait that made them highly desirable to peasant farmers—a colony’s ability to exploit floral resources over an area of at least 30 square miles (80 km2) [17]. This trait meant that the bee-keeper could exploit the production from land which he did not own (as beekeepers typically continue to do to this day).
Practical application: early “beekeepers” needed only to provide artificial nest cavities in areas where natural cavities had become scarce. The bees otherwise took care of themselves—foraging far and wide, and voluntarily returning home with the goods.
The practice of bee-keeping appears to have began in the Middle East, and then spread to other regions [18]. Early beekeepers, depending upon materials at hand, created all sorts of nest cavities (hives), such as horizontal or vertical hollow logs, clay pots or tubes, or straw skeps [19]. Horizontal hives were the norm in desert and Mediterranean climes; log gums, vertical hives, and skeps were often used in northerly (cold winter) regions. Once humans controlled the nest sites of the honey bee, thus began…
The Domestication Of The Honey Bee
Domestication: adaptation to intimate association with human beings.
Primitive beekeeping was not much different from predatory honey hunting, other than the hunters providing homes for their eventual prey within which to store the precious honey. So long as early bee-keepers practiced destructive harvesting (killing the colony in order to consume both brood and honey combs), little selective breeding would likely take place, due to the temptation to harvest the the most productive hives.
Clever beekeepers in the Mediterranean region (especially in the clay-rich Fertile Crescent) got around this problem by using horizontal clay tubes as hives, with the entrances to the front, and a removable plug in the back. Since bees tend to store honey away from the entrance, these beekeepers could harvest honey from the rear with a minimum of stinging by smoking the bees off the honey combs, without disturbing the broodnest. What a concept! Instead of killing the colony, one could “milk” it. (These tube hives were especially amenable to this practice, prior to the invention of movable frame hives. However, nondestructive harvest methods were also invented by “forest” and skep beekeepers [20] in northern regions).
The next thing they learned was how to propagate new colonies by transferring combs of brood and scoops of bees to new hives. They even learned how to transfer queen cells and virgin queens.
Practical application: this control of the queens meant that these beekeepers could then practice selective breeding, the foundation of the process of domesticating a species. I’m surprised by how few modern day beekeepers in this country selectively breed their own locally-adapted stock (since these “primitive” beekeepers were doing it 3000 years ago!).
Domestication is a sort of symbiotic mutualism, in which both the humans and the selected animals benefit. Beekeepers would certainly select for propagation those colonies that were most productive and amenable to being worked. Milner [21] explains:
The gentle behaviour of the major races of honey bee may be due, of course, to selection for this quality over many generations; even the “skep” beekeepers of former days would, no doubt, tend to destroy the worst tempered bees and retain the gentler colonies.
Not only would beekeepers select from local stock, but even import more desirable stock. Three thousand years ago, in the ancient city of Tel Rehov in Israel, commercial beekeeping was practiced [22] using a gentle, productive strain of bees imported from Turkey!
Limiting factor: Competition for food
Let’s suppose that beekeepers have now increased the available supply of skeps, gums, or Langstroth hives until the bee population is no longer limited by the number of nesting sites, but by something else? And now we get to the meat of the issue—competition for food resources. There is a limit to the number of colonies of bees that any area, no matter how rich in flowering plants, can support. That limit is called the carrying capacity of the landscape, and is commonly used to calculate how many livestock a pasture can support.
Beekeepers in my neck of the woods would no more brag about how much honey they made in a particular location than would a fisherman brag about the location of his favorite fishing hole. Should one do so, he’d likely find hundreds of new hives sitting on top of him the next season. This would be a perfect example of Garret Hardin’s influential concept of The Tragedy of the Commons [23], in which he points out that it may be to the individual herder’s benefit to add yet one more head of livestock to the common pasture, but to the herder community’s detriment once the addition of another animal exceeds the carrying capacity of the land (beekeepers today in some jurisdictions have wisely (and self-protectively) mitigated this inherent and inevitable problem by limiting commercial apiaries to registered locations, typically no closer than two miles apart) (Fig. 4).
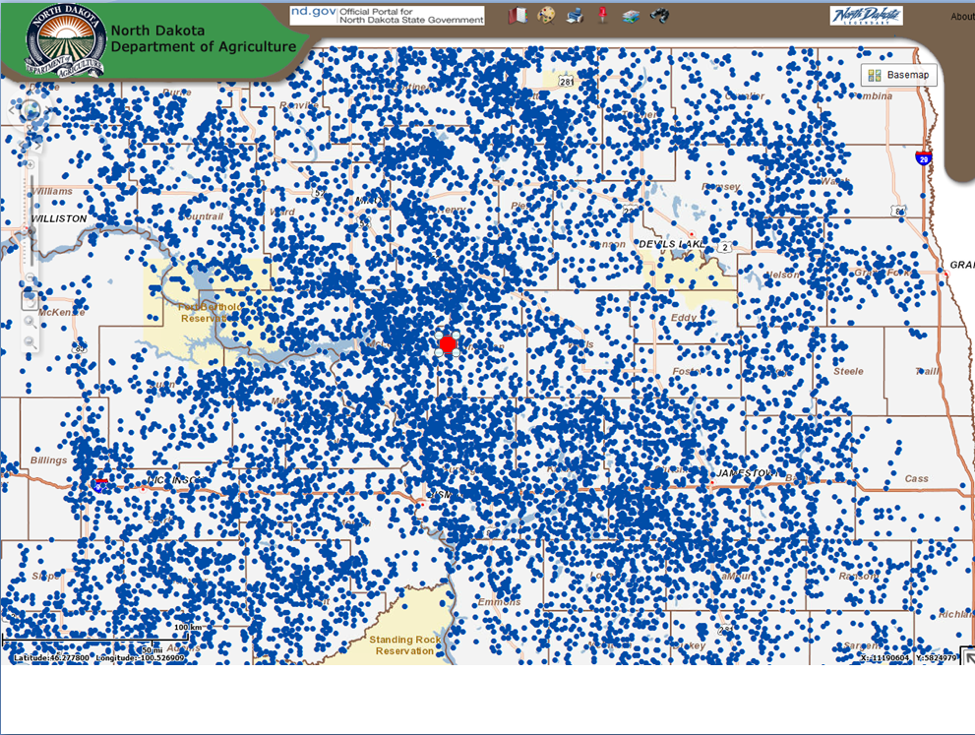
Figure 4. The Tragedy of the Commons exemplified. Locations of registered apiaries (blue dots) in North Dakota [[i]]. I added a 2½-mile-radius red circle in the center to indicate the area covered by the typical foraging range of a colony. Clearly, the forage areas of many of these locations overlap.
[i] https://apps.nd.gov/ndda/mapping/
And what sort of carrying capacities will various landscape types support? Studies have found natural colony densities of from 1–25 per square mile (the lesser density typical in temperate forests; the higher density in tropical areas, esp. with Africanized bees) [25]. Beekeepers of managed hives generally limit their apiary sizes so as not to exceed the carrying capacity of the land to produce a surplus honey crop.
Practical application: for example, in good forage areas of Montana and the Dakotas, beekeepers try to keep commercial apiaries a minimum of 2 miles apart. If such apiaries were placed on a 2-mi grid, that would allow 4 sq. mi. of forage area per apiary. Even at a high stocking rate of 48 hives per apiary, the stocking density would be only 12 managed hives per sq. mi. (6/mi2 at 24 hives per yard).
How does the density of managed hives in the European bee’s native range compare? Eva Crane cites records of hive density in Hungary in the late 1700’s of 30 to 460 per square mile! In the European Union today, in which beekeepers in some areas are complaining of poor colony performance, there are some 15 million reported managed hives, which works out to nearly 9 hives per square mile (perhaps exceeding the natural carrying capacity of the land).
By comparison, in the U.S. (which contains roughly the same percentage of arable land) the average density is only about 1 hive per square mile. Of course, average density over a continent does not reflect the actual hive loading of any particular area. Especially in the U.S., hives tend to be moved around, as opposed to the often stationary apiaries in Europe, and during summer, over half of all hives are located in only six states, accounting for only 16% of the U.S. land mass. However, even in those six states, the density of hives only starts to approach that of Europe as a whole!
Practical application: The Tragedy of the Commons definitely applies to beekeeping, since bees can’t be fenced in. I was driven out of a very good area in Nevada by beekeepers who moved in thousands of hives to the extent that I could hit another apiary with a thrown stone from any of my long-time locations. California has also reached that point in many areas, as beekeepers step on each others’ toes looking for any favorable place to place hives.
When I hear of all the bee problems in Europe, I wonder as to how much beekeepers there have contributed to the problem by overstocking hives on the available pasture. I’ll return later to the impact of modern agricultural practices upon the carrying capacity of agricultural land for bees.
Limiting factor: Reproductive success rate
In the absence of natural nest cavities, the survival of the honey bee depended in many areas upon the provision of such nest sites by humans, in the form of some sort of managed “hives.” And that fact gave beekeepers control over the reproductive success of any particular colony. By choosing which colonies were allowed to reproduce, beekeepers would rather quickly have been able to domesticate the bee by providing nest cavities only to those most amendable to husbandry. It would only have been in the scattered relict forests that wild, unmanaged populations of bees would have been able to survive. And this finally brings us to what I suspect is a major factor negatively affecting honey bees today:
The Price Of Domestication
Are honey bees truly a domesticated animal? And if so, how has that favored or hurt them? I’m out of space for now, but it gets more interesting…
A Note And Acknowledgements
Although I’ve spent considerable time in researching this article, my interpretation of the evidence is largely speculative. If anyone can add to this subject, please let me know.
As always, I am greatly indebted to my colleague Peter Borst, without whose research assistance I could not write these articles. And I cannot express how much I grateful I am for the words of appreciation from beekeepers worldwide, as well as their donations that support my research, writing, and website maintenance.
Citations and Footnotes
1 Ulrich Kotthoff, U, et al (2013) Greater past disparity and diversity hints at ancient migrations of European honey bee lineages into Africa and Asia. Journal of Biogeography 40: 1832–1838. Open access.
2 Adams, J (n.d.) Europe during the last 150,000 years. http://www.esd.ornl.gov/projects/qen/nercEUROPE.html This is a fascinating compilation of information and maps on the changes in climate and vegetation of Europe over time.
Ruttner, F (1988) Biogeography and taxonomy of honeybees. Springer-Verlag.
Miguel, I (2007) Gene flow within the M evolutionary lineage of Apis mellifera: role of the Pyrenees, isolation by distance and post-glacial re-colonization routes in the western Europe. Apidologie 38: 141–155. Open access.
Franck, P, et al (1998) The origin of West European subspecies of honeybees (Apis mellifera): New insights from microsatellite and mitochondrial data. Evolution 52(4): 1119-1134. Open access.
3 Delaney, D (2008) Genetic characterization of U.S. honey bee populations. PhD Thesis http://www.dissertations.wsu.edu/Dissertations/Summer2008/d_delaney_070108.pdf
4 The classification of these races is rather arbitrary, and under debate by taxonomists, but gives a general idea. There are also discrete breeding populations within each of the subspecies. See:
Radloff, SE, and HR Hepburn (1998) The matter of sampling distance and confidence levels in the subspecific classification of honeybees, Apis mellifera L. Apidologie 29: 491-501. Open access.
Al-Ghamdi, AA, et al (2013) Geographical distribution and population variation of Apis mellifera jemenitica Ruttner. Journal of Apicultural Research 52(3): 124-133. Open access.
5 http://en.wikipedia.org/wiki/Mediterranean_climate
6 Williams, M (2000) Dark ages and dark areas: global deforestation in the deep past. Journal of Historical Geography 26: 28–46. http://geography.fullerton.edu/taylor/ENST595T/darkages.pdf
http://www.fao.org/docrep/004/ab580e/AB580E02.htm
Oosthoek, KJW (n.d.) The Role of Wood in World History. http://www.eh-resources.org/wood.html
7 Milich, L (1997) Desertification. http://ag.arizona.edu/~lmilich/desclim.html.
8 Map from Adams, op. cit.
9 Adams, J, op. cit.
10 Walker, PA, et al (2003) Landscape changes in Nevada County reflect social and ecological transitions. California Agriculture 57(4): 115-121.
11 Seeley, TD and RA Morse (1978) Nest site selection by the honey bee, Apis mellifera. Insectes Sociaux 25(4): 323-337.
12 Thanks to Jay Miller and Darren Cox.
13 Johnson, S (1998) Who Moved My Cheese? G.P. Putnam’s Sons.
14 http://www.balqees.com/shop/yemeni/
15 (Broken Link!) http://www.yementimes.com/en/1633/business/1736/Liquid-pot-of-gold-Yemen’s-honey-trade.htm
16 http://www.thenational.ae/business/industry-insights/the-life/sticky-patch-for-yemen-honey-exports
17 Beekman, M and F Ratnieks (2000) Long-range foraging by the honey-bee, Apis mellifera. Functional Ecology 14(4): 490–496.
18 I’ve taken much of this historical information from Crane, Eva (1999) The World History of Beekeeping and Honey Hunting. Taylor and Francis Group.
19 I highly recommend the book The Quest for the Perfect Hive by Gene Kritsky (2010) Oxford University Press.
20 “Forest beekeeping” was practiced in Northern Europe, whereby beekeepers, would cut a recloseable door to a hollow high in a tree, allowing for repeated harvest without killing the colony.
21 Milner, A (2008) An introduction to understanding honeybees, their origins, evolution and diversity. http://www.bibba.com/origins_milner.php This is an excellent review of the domesticated races of the honey bee, and a well-thought plea for the breeding of locally-adapted stocks. See also:
(Broken Link!) http://www.aragriculture.org/insects/bees/races.htm
22 Mazar, A and N Panitz-Cohen (2007) It is the land of honey: Beekeeping at Tel Rehov. Published in Near Eastern Archaeology 70(4): 202-219. Open access.
Bloch, G, et al (2010) Industrial apiculture in the Jordan Valley during Biblical times with Anatolian bees. PNAS 107(25): 11240-11244. Open access.
23 Hardin, G (1968) The Tragedy of the Commons. Science 162(3859): 1243-1248. http://www.sciencemag.org/content/162/3859/1243.full
24 https://apps.nd.gov/ndda/mapping/
25 Ratnieks, FLW, et al (1991) The natural nest and nest density of the Africanized honey bee (Hymenoptera, Apidae) near Tapachula, Chiapas, Mexico. Can. Entomol. 123: 353-359.
Baum, KA, et al (2005) Spatial and Temporal Distribution and Nest Site Characteristics of Feral Honey Bee (Hymenoptera: Apidae) Colonies in a Coastal Prairie Landscape. Environmental Entomology 34(3):610-618.
Taber, S, III (1979) A population of feral honey bee colonies. Am. Bee J.ABJ 119: 842-847.
First published in: American Bee Journal, March 2014
CONTENTS
We Are Two Adaptive Species
Honey Bees Are Designed For Rapid Adaptation
The Human-Facilitated Realized Niche Of The Bee
The Natural Limiting Factors Of The Honey Bee Population
Limiting Factor: The Weather
Limiting Factor: Predation
Limiting Factor: Competition For Nest Cavities
Limiting Factor: Carrying Capacity Of The Habitat
Limiting Factor: Reproductive Success Rate
Limiting Factor: Competition And Intercolony Parasitism
Limiting Factor: Pathogen Transmission
A Special Case: The Viruses
Conclusion
Footnotes and Citations
What’s Happening to the Bees?
Part 2
Originally published in ABJ March 2014
Randy Oliver
ScientificBeekeeping.com
Over the past few decades, as a beekeeper/biologist I’ve had the opportunity to watch evolution in action. I’ve observed the catastrophic effects upon colony health due to the introduction of new parasites, periodic pathogen epidemics, and the more subtle effects of changing land use practices and climate change. I’ve also witnessed the evolution of both recreational beekeeping and the bee industry, as we’ve been forced to change our management practices and income streams due to the aforementioned biological and environmental factors, plus changes in markets and the impacts of regulatory decisions.
My point is that things change, each change altering the realized niche of both bees and beekeepers. Neither Nature nor the Market are the least bit sentimental. Those who don’t adapt to change die.
The adaptive process may make headlines, or it may go largely unnoticed. But one thing’s for sure—the more that we understand the changes in the parameters of the bee and beekeeper niches, the better we can successfully engage in the adaptive process.
We Are Two Adaptive Species
Both Apis mellifera, and those Homo sapiens categorized as “beekeepers” have proven to be incredibly adaptive species (ecologists would say that we exhibit a high degree of plasticity). At times, each of our populations face limiting factors that weed out the least fit. And the higher the failure rate of individual colonies or beekeeping operations, the stronger the selective pressure to adapt.
So what sort of failure rate is “normal” for bees under optimal natural conditions (the fundamental niche)? Let’s do the math! Given: an established and stable local population of bees under optimal conditions. In the spring each colony will produce at least one swarm. At that point, the colony population will have temporarily at least doubled. But such a rate of reproductive increase is obviously unsustainable, since by definition a “stable” population ends with the same number of colonies each year. So simple arithmetic tells us that in nature, on average, at least half of all colonies will succumb each season, even under the best of conditions.
Although half the colonies will fail on average, it is not the average colony that fails. The key point is that it is the least fit that tend to fail, and a greater proportion of the most fit survive. This considerable amount of selective pressure is what drives adaptation and evolution in each natural population of bees.
It’s the same with beekeepers. The failure rate for beginning beekeepers is often even higher than the above. Commercial operations also fail; despite record high prices for honey and pollination services, our recent elevated colony mortality rate is eliminating the profit margin for some operators, who, sadly, will go out of business. But, again, the hard fact is that it is the less fit (or less adaptable) operations that are failing—I speak with plenty of operators who aren’t experiencing egregiously high winter losses, and whose businesses are firmly in the black. It is these operations that are successfully adapting to their realized niche.
Honey Bees Are Designed For Rapid Adaptation
As I study the honey bee reproductive strategy, one principle jumps out:
Everything about the honey bee reproductive process is designed for rapid adaptation to changes in its realized niche, and for the recovery of the bee population from decimation events [1].
Apis mellifera has the highest known rate of genetic recombination of any animal. And those “experimental” recombinations are then filtered for success through the haploid drones, who have only one set of chromosomes, meaning that only the best novel combinations of genes (alleles) have any chance of being passed to the next generation.
The genetic combinations of the fittest drones then predominate in the matings that occur in drone congregation areas, thus ensuring that each virgin queen has the best chance of loading up with the best genetics that the overall bee population in that area has to offer. And she further ensures the maximum diversity of her offspring by mating with multiple drones. Add to this the incredible epigenetic plasticity of the honey bee [2], and we have an organism able to quickly adapt to whatever Nature throws at it!
I observe something similar with beekeepers. Those who are consistently trying new things and adapting to the changing biological and business environments are those who tend to be the most successful in the long run.
The Human-Facilitated Realized Niche Of The Bee
OK, so in nature, half the colonies fail each year. But provided facilitation by human beekeepers, that rate can drop to around 10% for well-managed operations. However, in recent years, U.S. beekeepers have been reporting distressingly high rates of colony failure. Clearly, something in the realized niche of our honey bees has changed in the last decade (one or more limiting factors). In order to attempt to figure out exactly which factors are responsible, let’s first determine what the primary limiting factors were for honey bee populations prior to humans. Perhaps then we can better understand how our actions (and the Earth’s burgeoning human population) are affecting honey bee survival. And maybe then we can take steps to make life easier for our beloved bees (and improve the bottom line of those of us who make our living at keeping ‘em).
The Natural Limiting Factors Of The Honey Bee Population
Limiting Factor: The Weather
The weather would be an obvious limiting factor—colonies are stressed by extreme cold, unfavorable flight weather during the spring or summer, or by lack of forage and water during droughts. Such intermittent weather events may sporadically cull the bee population, but would only affect long-term adaptation if they occurred regularly. So let’s focus upon which factors limited bee populations in favorable years, during which they have the intrinsic ability to increase exponentially.
Limiting Factor: Predation
Honey bees are herbivores. The populations of many herbivores are controlled by predators (Fig. 1). If the population of the prey species becomes dense, predator species ramp up their numbers to take advantage of the food source. The result is typically an oscillating predator/prey population dynamic (this is one of the bases for integrated pest management in agriculture).

Figure 1. Foraging bees face plenty of predators, such as wasps, assassin bugs, birds, spiders, and dragonflies. One of “my” bees (above) became a meal for a robber fly.
Although predators of foraging bees may take a bite out of the population of older bees in the hive, they do not appear to be a primary limiting factor of the overall bee population. An exception to this rule might be the Asian Hornet (Vespa velutina) (now introduced in Europe), which can decimate small colonies of bees by picking off returning foragers [3].
A more serious form of predation is direct invasion of the hive. A healthy colony can generally repel or otherwise deal with small invaders such as ants, wasps, and Small Hive Beetles. More problematic are those large mammalian predators willing to ignore the bees’ defensive stinging, such as bears, skunks, honey badgers, and humans. The bees’ main defense against such predators is to nest in inaccessible fortifications. And that leads us to…
Limiting Factor: Competition For Nest Cavities
Bees are pretty picky about the nest cavities that they choose, strongly preferring elevated tree cavities having small, defendable entrances [4] (Fig. 2). In treeless areas, the lack of suitable nest sites could well have been a limiting factor for the honey bee population. But this is unlikely to have been the primary limiting factor anywhere that patches of ancient forest were within range.

Figure 2. This hollow black oak provides a protected nest cavity impenetrable by our local black bears.
Although a lack of suitable cavities may not the main limiting factor of the bee population, it does bring to mind another fascinating aspect of bee behavior. Please allow me to digress for a bit. Not all beekeepers are aware that intra- and inter-species parasitism is common among bees, wasps, and ants. For example, queens of a number of bumblebee species parasititically invade and take over other bumblebee colonies. And the Cape Bee (Apis mellifera capensis) is famous for its ability to parasitically take over colonies of the Savannah Bee (Apis mellifera scutellata) [5]. This sort of deplorable behavior appears to be ingrained in the bee genome– bees covet the fruits of their neighbors’ hard work.
Part of the Africanized honey bees’ ability to rapidly expand its range in the Americas was likely its ability to invade and usurp the established nests of European bees [6]. Lately, Dr. Wyatt Mangum has been reporting on his observations of similar behavior by ostensibly non-Africanized bees in Virginia [7].
Such takeovers give the usurping swarm a profound advantage. Rather than needing to establish a nest and provision it with stores from scratch, it can simply take over an established colony, essentially hijacking its combs, stores, and entire workforce. Although Mangum, so far as I know, is the first to report such behavior for European honey bees, his observation may answer a question that has been bugging me for years: why do European bees send out what appear to be doomed swarms in late summer?
Bees typically swarm in spring, for the obvious reason that that timing allows the swarm colony to establish and provision a nest in time for winter. But in actuality, there are two peaks of swarming during a season (Fig. 3). I’d long noticed this, and wondered why in the heck a colony would bother to swarm in the late summer—natural selection should have eliminated such suicidal behavior. It just didn’t make sense!

Figure 3. Seasonal timing of swarm emergence dates in New York, showing the main late-spring peak, followed by a lesser peak around the first of September. Why the heck would colonies swarm in late September? After Seeley [[i]].
[i] I transcribed and plotted the data from Fig. 1 in Seeley, TD, et al (1989) Bait hives for honey bees. Cornell Coop Ext Inf. Bull. No. 187. http://ecommons.cornell.edu/bitstream/1813/2653/2/Bait%20Hives%20for%20Honey%20Bees.pdf
I assumed that the data was for New York and from the following paper, but I was unable to obtain a copy:
Fell, R. D., et al (1977) The seasonal cycle of swarming in honeybees. J. Apic. Res. 16:170-173.
But Mangum’s observations reminded me that I’ve also seen in past years late-summer swarms landing on hives, and also seen balled queens in hives in late summer. Perhaps I simply never put it all together! Could it be that this is an innate, but previously unrecognized, behavior in European bees?
It is certainly biologically plausible that under certain circumstances European colonies behave like their Africanized brethren (they are, after all, the same species), and send out older queens who have already successfully built up a colony (and perhaps even a second swarm colony), but still have enough vigor left to do an invasive usurpation of a nearby nest. If the colony were already superseding that queen, then it would be an inexpensive gamble to send her out, accompanied by a hit team of experienced workers, to try to take over an unsuspecting colony, its stores, and its workforce shortly before winter.
Pardon my digression—let’s return to our search for the main limiting factor of the natural bee population.
Limiting Factor: Carrying Capacity Of The Habitat
The maximum population density for the realized niche of a population is set by the carrying capacity of that particular environment, typically limited by resources such as food. However, the honey bee is a special case; similar to the bear, the colony can gorge when food is plentiful, and store “fat” (honey and beebread) as reserves for lean times (as during overwintering).
There are indeed areas in which colonies can barely put on enough honey during the main flow in order to make it through the winter. But by definition, such areas would not meet a primary requirement of the fundamental (optimum) niche of the honey bee, so we can disregard such areas from this discussion. What we are interested in is the limiting factor of the bee population in areas that normally produce a good honey flow.
Limiting Factor: The Timing Of The Bloom
Honey bees are defined by their ability to store food reserves—honey and beebread—to see them through lean times. But there are times other than the main honey flow during which the availability of nectar, and especially pollen, are of critical importance to the ability of colonies not only to survive, but also to reproduce. Colonies must build up and produce a swarm early enough in the season that both parent and swarm colony have fighting chances to store enough honey to make it through the following winter [9]. Such buildup requires the initiation of broodrearing in the middle of winter, which is in turn dependent upon having stored a large supply of beebread during the fall pollen flow [10].
What we must keep in mind is that a colony of bees is only effective at putting away a honey surplus if it has grown a large enough population to efficiently forage upon and store the available nectar. Timing is everything. Too large a population at the wrong time of the year would be counterproductive, since those hungry mouths would consume more honey than they were able to store. A locally-adapted population of bees times its buildup to coincide with the main flow, and then quickly shrinks back to survival mode.
Many of us tend to base our idea of typical bee behavior upon that of commercially-selected Italian stock. Such stock, originally adapted to Mediterranean climates, and bred for continuous broodrearing and high honey production, can hardly be expected to be representative of wild type bees adapted to cold-winter areas (I am not dissing Italian-type bees—they are well adapted to build up early for almond pollination).
Bees adapted to colder winters, such as the Carniolans or Russians, are far more responsive to the environment, especially to the availability of pollen. As soon as plants start producing pollen in spring, bees of these races explode into action —working even in cool and wet weather, and madly brood up.
The reason for their frenzy is that they must build up their population early enough to produce a swarm in time for it to have a decent chance at establishing a nest and putting away adequate winter honey stores during the brief main honeyflow (typically May through June; in temperate climates, colonies may only gain weight for a few weeks a year). I’ve previously graphed the bee colony’s amazing ability to quickly build up [11]. An absolute requisite for such a rapid rate of growth is a monster supply of pollen in the early spring.
After the main flow comes the summer dearth. When pollen is unavailable, wild-type bees sit tight in survival mode [12], cooling their heels and conserving their energy. I’ve seen Russian bees cease broodrearing in August in the arid California foothills, appearing as though they were queenless. In the adjacent yards, my Italian stock just keep on rearing brood, and required supplemental feeding of protein in order to keep them in decent shape for going into winter..
So I suspect that the limiting factor for bees in a natural realized niche is not the amount of food available during a brief period of food abundance (the main honey flow), but rather the quality and quantity of forage available during spring and late summer/fall.
Practical application: the realized niche of the honey bee is likely largely defined not by the amount of nectar available during the main honey flow, but rather by the quantity and quality of pollen available prior to and after that period. A successful colony requires a dependable abundance of quality pollen and nectar for early spring buildup; and then adequate late-season pollen to ensure its ability to produce a cluster of protein-rich “winter bees” and to store beebread for midwinter broodrearing.
Nevertheless, prior to man, biologically productive areas typically supported a diversity of native vegetation that produced exactly such an extended season-long buffet of nutrition. Thus, I doubt that during favorable years, a lack of available spring or fall food resources was the limiting factor of the bee population prior to man.
Limiting Factor: Competition For Those Food Resources
Competition for food resources, either against other species or one’s own species, is a common limiting factor of the realized niche. So what sort of competition do bees face?
Anywhere that there are flowering plants, there are pollinators that have coevolved with them. Most are insects (although in some areas, birds, bats, or other mammals may be important). Does competition with other pollinators limit the honey bee population?
Let’s think about it. Since one can place a hive of bees into most any favorable habitat and still make honey despite the presence of established populations of native pollinators, I suspect that competition with other species is not normally the limiting factor of the honey bee population. On the other hand, as any beekeeper quickly recognizes, honey bee colonies certainly compete with one another!
Beekeepers tend to focus upon the amount of nectar available during the main honey flow, and understand that one can overload an area with managed hives. But how about the density of a natural population of bees—does the amount of nectar available during the main flow limit that population?
Again, let’s check easily-verifiable observations. In areas with well-established populations of feral bees (Australia, Hawaii, formerly in the U.S), one can bring in additional managed colonies, yet still produce a substantial honey crop during the main flow—clearly, nectar is produced in excess of what an established natural population of bees can harvest. It follows then that at a “normal” population density of unmanaged colonies, competition for nectar during the main flow was not the limiting factor for colony survival.
On the other hand, competition for pollen during spring or fall could well be a limiting factor. And along with that competition comes…
Limiting Factor: Intercolony Parasitism
Competition between colonies does not occur only for nest sites or at the flower; it can also happen directly at the hive. All’s fair in love, war, and in Nature. If you can save yourself effort by stealing the fruits of another’s labor, so be it. The term for this is kleptoparasitism (kleptoparasite: a bird, insect, or other animal that habitually robs animals of other species of food).
What we call “robbing” is a form of kleptoparasitism, and during early spring or the summer dearth, the robbing pressure between colonies can brutal. There would be a clear competitive advantage to those colonies that successfully robbed honey from others; conversely, there would be an advantage to those colonies best able to defend their hard-won stores.
Robbing behavior may also be more insidious than the overt invasion of a weak colony by a strong colony. Dr. Wyatt Mangum detailed the sneaky “progressive robbing” of one colony by another [13]. Such robbing would constitute an insidious drain upon the victim colony. In areas of high colony population density, I suspect that robbing pressure–at times other than during major honey flows–is a limiting factor in colony density.
And this very robbing behavior brings us to our last suspect factor– that famous Horseman of the Apocolypse, Pestilence.
Limiting Factor: Pathogen Transmission
As most beekeepers soon find out, honey bees are host to a number of parasites and pathogens. To a biologist, a pathogen is a parasite (such as a virus, fungus, or bacterium) that can cause disease. These parasites can cause the bees to suffer from either endemic or epidemic infections. A well-adapted parasite typically does not, under normal circumstances, cause serious disease, but rather smolders in the bee population of each individual colony as an endemic infection; sacbrood virus or Nosema apis follow this model. These well-adapted parasites are typically vertically transmitted, that is, from parent to offspring, or in the case of bees, from mother colony to daughter colony. They can generally be found in a colony, but so long as the colony is not stressed, there are no symptoms of disease.
Other parasites tend to go epidemic, sometimes in recurring cycles; chalkbrood, Chronic Bee Paralysis Virus, American Foulbrood, and European foulbrood fall into this category. Individual colonies are able to “clear” themselves of these parasites; the parasites maintain a presence in the local bee population, but may not be found in every colony. Epidemic parasites are largely dependent upon horizontal transmission from colony to colony, and unlike the well-adapted endemic parasites, actually benefit by the weakening or death of an infected colony. As such, they would be considered to be density dependent infectious diseases. As the density of the host (honey bee colonies) in the environment increases, the opportunities for transmission of the parasite from colony to colony directly increases [14]. The pathogen causing the disease can only persist if the host density exceeds a certain threshold (if bee colonies were scattered beyond flight distance, there would be slim possibility for an infectious pathogen to transmit from one colony to another).
Conversely, not being regularly exposed to a pathogen removes the selective pressure for the bees to maintain genetic (or epigenetic) resistance. Thus, in nature, epidemics of certain pathogens ebb and flow, often decimating a host population one year, at which point the host density is decreased to the extent that the pathogen nearly disappears, struggling to maintain a foothold in the few remaining, and most resistant, hosts. As the host population then recovers over the years, an epidemic may then recur (higher host densities favor virulent mutations of the pathogen). This is a common cyclic pattern in insect species with large populations, and bees are no exception [15].
Practical application: After a plague it may take years for a particular pathogen to again recover its hold in the bee population. And it may take a special combination of environmental circumstances, and perhaps coinfection with other parasites, in order for it to do so.
For most species of wildlife, natural populations tend to reach some sort of dynamic equilibrium, with pestilence being the ultimate limiting factor if all other conditions are optimal. Anderson [16] explains:
It is likely that interplay between the pathogenicity of viral, bacterial, [or] protozoan infections and the nutritional state of the host contributes importantly to the density-dependent regulation of natural populations, with the parasites greatly amplifying the effects of low levels of nutrition.
Such pestilence typically occurs in the form of epidemics, during which the pathogen(s) efficiently spread through a stressed and overcrowded population, the key word being overcrowded. Too many colonies of bees in an area is a recipe for disaster—a ticking time bomb just waiting for the right combination of environmental circumstances and the presence of a virulent pathogen (or combination thereof).
A Special Case: The Viruses
But it’s a bit more complicated when reservoir hosts are involved. That is, when a pathogen is not limited to honey bees as its sole host. And this is the case with the “bee” viruses, most or all of which appear to actually be generic insect viruses that bees pick up when they visit flowers. So it is likely that the density of the bee population is not limited merely by diseases specific to honey bees, but also by the entire pool of viruses that infect pollinating insects [17] (Fig. 4).
Breaking news: As I type these words, a collaboration of researchers associated with the USDA ARS labs is about to release a stunning paper [18], in which they detail how a plant virus is now infecting both honey bees and varroa, and appears to be associated with collapsing colonies. Their findings suggest that varroa may be a key player in the cross-kingdom jump of this virus. The complexity of the bee, mite, plant, and virus web of infection continues to astound us!

Figure 4. These Russian bees near the hive entrance are in the process of cooking a too-bold bald-faced hornet to death [[i]]. Hornets and yellowjackets eat bees, and are thus exposed to bee pathogens. Exposure also goes the other way as the wasps contaminate nectar with virus particles when they visit flowers, or, as in this case, if the dying wasp exudes any body fluids.
[i] Ugajin A, et al. (2012) Detection of neural activity in the brains of Japanese honeybee workers during the formation of a “hot defensive bee ball”. PLoS ONE 7(3): e32902.
Historically (meaning prior to varroa), these viruses were sporadically present in bee colonies, but generally as inapparent (free of noticeable symptoms) infections [20]. It was only under certain circumstances that they went epidemic and caused noticeable morbidity or mortality of colonies:
Taken together, these data indicate that bee virus infections occur persistently in bee populations despite the lack of clinical signs, suggesting that colony disease outbreaks might result from environmental factors that lead to activation of viral replication in bees [21].
In any case, in some years in some localities, such “perfect storms” of environmental factors, virulent mutations of one or more pollinator viruses, and coinfections with other parasites have historically led to serious colony collapse events [22]. This is not a new thing!
Conclusion
My original question was what were the primary limiting factors in the realized niches of the honey bee prior to human influence? I hope that I have adequately covered these factors, since I feel that our understanding of them is critical for us to be better beekeepers, and to make good management decisions.
As always, I prefer to let the reader draw his/her own conclusions, but to me it appears that that the primary limiting factors in colony survival in favorable areas were most likely the density-dependent competition for pollen during spring and fall, coupled with the associated transmission of certain pathogens.
The above factors have long been associated with epidemics in the bee population. It appears to me that our bees today are in the midst of an ongoing and complex multi-pathogen epidemic largely precipitated by the actions of mankind. In the next installment of this article I will explore how this situation came about, examining how changes in world trade, agriculture, the environment, and in beekeeping practices have affected the realized niche of the bee, its parasites and pathogens, and the business models of beekeepers. My hope is that by fully understanding how we inadvertently helped to create the problem, that perhaps we can better take steps to help our poor bees deal with the problem, and for ourselves to stay in business in the process.
Footnotes and Citations
1 “Decimation events” being plagues, droughts, wildfires, extreme weather events, etc.
2 http://www.nature.com/news/job-swapping-makes-its-mark-on-honeybee-dna-1.11418#/b1
3 Tan, K, et al (2007) Bee-hawking by the wasp, Vespa velutina, on the honeybees Apis cerana and A. mellifera. Naturwissenschaften 94(6): 469-472. Open access.
4 Seeley TD and RA Morse (1978) Nest site selection by the honey bee, Apis mellifera. Insectes Sociaux 25: 323–337.
5 Martin, S, et al (2002) Usurpation of African Apis mellifera scutellata colonies by parasitic Apis mellifera capensis workers. Apidologie 33: 215-232.
Danka, RG, RL Hellmich, TE Rinderer (1992) Nest usurpation, supersedure and
colony failure contribute to Africanization of commercially managed European honey
bees in Venezuela. Journal of Apicultural Research 31 (3/4): 119-123.
6 Schneider, SS, et al (2004) Seasonal nest usurpation of European colonies by African swarms in Arizona, USA. Insectes Sociaux 51(4):359-364.
7 Mangum, W (2010) The usurpation (takeover) of established colonies by summer swarms in Virginia. ABJ 150(12): 1139-1144.
Mangum, W (2012) Colony takeovers (usurpations) by summer swarms: they chose poorly. ABJ 153(1): 73-75.
Mangum, W (2013) Summer swarms with queen balling. ABJ 153(2): 163-165.
8 I transcribed and plotted the data from Fig. 1 in Seeley, TD, et al (1989) Bait hives for honey bees. Cornell Coop Ext Inf. Bull. No. 187. http://ecommons.cornell.edu/bitstream/1813/2653/2/Bait%20Hives%20for%20Honey%20Bees.pdf
I assumed that the data was for New York and from the following paper, but I was unable to obtain a copy:
Fell, R. D., et al (1977) The seasonal cycle of swarming in honeybees. J. Apic. Res. 16:170-173.
9 Otis, GW and JM Wearing-Wilde (1992) Net reproductive rate of unmanaged honeybee colonies, (Apis mellifera L.). Ins. Soc. 39:157-165.
10 Seeley, TD and PK Visscher (1985) Survival of honeybees in cold climates: the critical timing of colony growth and reproduction. Ecological Entomology 10: 81-88.
11 https://scientificbeekeeping.com/sick-bees-part-17-nosema-the-smoldering-epidemic/
12 This physiological change to diutinus bees occurs when newly-emerged workers sense queen pheromone, but no young brood pheromone, which tells them that the colony is in survival mode. This typically occurs in fall, but can also occur during summer dearth. See https://scientificbeekeeping.com/an-adaptable-workforce/
13 Mangum, W (2012) Robbing: Part 2: Progressive robbing. ABJ 152(8): 761-764.
14 Hudson PJ, et al (2001) The Ecology of Wildlife Diseases. Oxford University Press, Oxford.
15 https://scientificbeekeeping.com/sick-bees-part-9-pathogens-and-plagues/
16 Anderson, RM and RM May (1979) Population biology of infectious diseases: Part I. Nature 280(2): 361-367.
17 Singh R, et al (2010) RNA viruses in hymenopteran pollinators: evidence of inter-taxa virus transmission via pollen and potential impact on non-Apis hymenopteran species. PLoS ONE 5(12):e14357.
18 Lian JL, Cornman RS, Evans JD, Pettis JS, Zhao Y, Murphy C, Peng WJ, Wu J, Hamilton M, Boncristiani HF, Jr., Zhou L, Hammond J, Chen YP. 2014. Systemic spread and propagation of a plant-pathogenic virus in European honeybees, Apis mellifera. mBio 5(1):e00898-13. doi:10.1128/mBio.00898-13. Open access.
19 Ugajin A, et al. (2012) Detection of neural activity in the brains of Japanese honeybee workers during the formation of a “hot defensive bee ball”. PLoS ONE 7(3): e32902.
20 Hornitzky, M (1987) Prevalence of virus infections of honeybees in eastern Australia. Journal of Apicultural Research 26(3) : 181-185.
Anderson, DL and AJ Gibbs (1988) Inapparent virus infections and their interactions in pupae of the honey bee (Apis mellifera Linnaeus) in Australia. J. Gen. Virol. 69: 1617-1625.
Mouret, C, et al (2013) Prevalence of 12 infectious agents in field colonies of 18 apiaries in western France. Revue Méd. Vét. 164(12): 577-582. http://www.revmedvet.com/2013/RMV164_577_582.pdf
21 Tentcheva, D, et al (2004) Prevalence and seasonal variations of six bee viruses in Apis mellifera L. and Varroa destructor mite populations in France. Appl. Environ. Microbiol 70(12): 7185-7191. http://aem.asm.org/content/70/12/7185.full
22 Underwood, R and D vanEngelsdorp (2007) Colony collapse disorder: have we seen this before? Bee Culture 135(7): 13-18. http://ento.psu.edu/pollinators/publications/underwood
First published in: American Bee Journal, February 2014
What’s Happening to the Bees?
Originally published in ABJ Feb 2014
Randy Oliver
ScientificBeekeeping.com
I’m realizing that what I thought was going to be a quick review of CCD has turned into a very long series of detailed articles, and I’m not even near reaching the conclusion. So on this 20th anniversary of my first seeing the parasite that changed beekeeping worldwide, I thought that I’d interrupt the Sick Bees series and attempt to explain, more briefly (hah!), and from an ecological perspective, what’s happening to the bees these days, and how beekeepers are being forced to adapt.
One Wild Ride!
In 1994 I saw my first varroa mite—on a stickyboard that had been placed by the county bee inspector under one of my hives. Little did I know how much that little maroon speck was about to change my life!
Varroa clobbered me (or more precisely my bees). And then did it again; and then again. I finally got pissed off and decided that I should either give up beekeeping altogether, or dust off my scientific training and really learn about bee and mite biology, and how to apply it to practical beekeeping.
What’s interesting is that my very first article (on almond pollination), in the fall of 2006, was published just as Dave Hackenberg’s colonies began to collapse from CCD. The occurrence of CCD put the bee research community into high gear to try to figure out what the heck was happening. And that created a “niche” for a practical beekeeper with a biological background, willing to act as a translator of the scientific findings to an alarmed beekeeping community. I unwittingly stepped into that niche, and it swallowed me up. A mere seven years later, and to my utter surprise, I’ve gone from being an obscure sideline beekeeper to a globetrotting speaker on bee health and management.
I’ve now been fortunate enough to visit both professional and recreational beekeepers in every part of North America and in several other countries. I’ve seen many styles of beekeeping, from the tropics to the Arctic, heard the local problems and concerns, and had the chance to learn from some very smart and successful beekeepers. I’ve attended scores of conferences, read countless scientific papers, and picked the brains of the world’s best apicultural researchers. Then I’ve done my best to share what I’ve learned with others. I’ve met scores of wonderful people and made a lot of new friends, and I’d like to take this opportunity to thank you all for the appreciative and effusive support!
How We’ve Benefitted From CCD
CCD has been a mixed blessing to beekeepers. It brought grown men and women to tears (see the film The Last Beekeeper [1]), and the elevated rate of colony mortality in recent years has made it difficult to keep our operations in the black. But it also pushed our scientific community to learn more about the biology of the honey bee than they had in a great many years. And many of us are much the better beekeepers for it.
Unlike that of other livestock, the true contribution of pollinators to U.S. agricultural production is not reflected by farm gate sales figures, so bees have traditionally not received their fair share of USDA research funding, nor does the beekeeping industry have the lobbying clout of the cattle, poultry, or pork producers. But we’ve benefitted from the public awareness of the plight of pollinators, which has resulted in the shifting of some funding our way [2] (although bees still only get about a tenth the amount of money set aside for research on beef production). In addition, universities, grantors, and other governments have recently supported a great deal of research into honey bee and pollinator health (I only wish that the millions who signed the internet petitions to “save the bees” had instead each donated a single dollar toward bee research).
Misunderstanding And Misinformation
Although CCD refers to a specific set of symptoms [3], the media soon began to use the term for any sort of honey bee mortality (as did many beekeepers). And although the epizootic appears to have largely run its course, speculation ran rampant as to the cause(s) of “CCD,” and continues to do so with every new “documentary” and press release. Although “CCD” remains the poster child of colony losses, a blue-ribbon group of bee researchers cautions:
During the winter of 2008/2009, ∼10% of the 2.3million managed honey bee colonies in the US died with “CCD-like symptoms”, and US beekeepers self-diagnosed CCD as only the 8th most important contributor to colony mortality, behind starvation, queen-related issues, and parasites. The point is, honey bees die from many things. We must be careful to not synonymize CCD with all honey bee losses [4] (emphasis mine).
I’m typing these words as I fly over the beautiful jigsaw-puzzle-like frozen Manitoba landscape on my return from a speaking engagement in Sweden, where the film More than Honey had been recently shown. To my considerable surprise, the Swedish beekeepers (Fig. 1), after viewing the movie, were under the very strong impression that the bee problems in the U.S. were due to our brutal commercial beekeeping practices, and the moving of hives to the deadly almond orchards in California.

Figure 1. Beekeeper Göran Sundström at one of his apiaries in Sweden. Göran typically keeps 12 hives in an apiary, and goes to considerable trouble to comply with some rather arbitrary rules to have his honey certified as “organic.” The red paint is a traditional color for rural buildings in this area.
As it happens, the director of that film had stayed with me during his initial scouting visit to the U.S., and I was responsible for introducing him to my friend John Miller, who was unfortunately (and I’m sure unknowingly) to be cast in the role of the evil bee abuser, so I felt some responsibility to dispel those misconceptions to the concerned audience. And this brings me to my next subject…
Bees are currently enjoying a great deal of attention from a fearful public eager to do something, anything to help them. This could be a really good thing for the bees, for beekeepers, and for the environment as a whole if such public concern and activism could be guided into meaningful actions. I can’t really blame the public for being confused, since the entertaining and sensational docu-dramas about the impending extinction of the honey bee resonate more emotionally than do the dry and qualified explanations by scientists as to the “multifactorial” causes of colony mortality.
As a result of all the misinformation and hysteria out there, an unsure and distrustful public puts pressure their representatives to pass this or that new regulation to “save the bees.” This scares me. I feel that we should heed the sage advice of Thomas Jefferson:
People are inherently capable of making proper judgments when they are properly informed.
And therein lies the problem: due to the complexity of what’s happening with bees these days from the biological, environmental, agricultural, and economic standpoints, it’s danged hard to be “properly informed.” My gosh, just look at me trying to do that “informing”—I first thought that the “Sick Bee” series was only going to be two or three articles long! So what to do?
Challenging One’s Beliefs
One should be careful about embracing the popular stories about why “the bees are dying.” Some of the myths resonate so emotionally that they win uncritical acceptance by the mainstream, despite the fact that they cannot be reconciled with obvious facts (e.g., that bees can indeed thrive surrounded by GMO corn and soy, or on neonic-treated canola). As the popular scientific author Stephen Jay Gould pointed out:
The most erroneous stories are those we think we know best – and therefore never scrutinize or question.
Anyone who knows me (or has had the misfortune of trying to promote an unsubstantiated argument in my presence) can tell you that I’m a challenging and provocative person by nature. I’ve found that the best way to get to the truth is to learn how to argue your opponent’s side of a debate as well as you can argue your own. Therefore, I am more than willing to play Devil’s Advocate any time that I see one side of a legitimate argument being ineffectually presented. And I’m ruthlessly skeptical of any claims that do not jibe with what I see with my own eyes.
As you can imagine, this has earned me my share of vitriol from those who “know the truth” (read [5] for an enlightening discussion). Luckily, my mailbox runs about a hundred to one with thank you letters from beekeepers who appreciate my evaluation of the issues. I take this responsibility seriously; in order to remain objective and unbiased, I go out of my way to constantly question every one of my opinions (I avoid making “conclusions”). I eschew holding any “beliefs,” but rather adhere to the following principles:
- That I should respect Nature and all forms of life (my ethical environmentalist side),
- That I should thoroughly investigate all research and explanations of any subject, and avoid cherry picking data that suits my ideological convictions (my curious open mindedness and willingness to do my homework).
- That I should base my opinions upon information and experimental results which stand up to scrutiny and questioning (my scientific side),
- That I should then truth-check those opinions against on-the-ground evidence and observations (my practical side).
Unfortunately, many crusaders allow their commendable environmental consciousness (and innate fear of technology) to override the last three principles, which is understandable, since doing the homework is really hard, and our understanding of the biology involved is as yet incomplete. But I have some suggestions as to where to start…
A Homework Assignment
Allow me to first assign you some required reading. Put down this article and read Berndt Heinrich’s fascinating book Bumblebee Economics [6]. Heinrich studied the minute details of exactly how bees make a living in their ecological niche, focusing upon the economics of energy utilization. His revelationary insights changed my understanding of bee life completely. Then for something entirely different read Ron Miksha’s Bad Beekeeping [7] for a perspective on the economic trials and tribulations faced by professional beekeepers (his comments on p. 243 are especially relevant). I’ll wait ‘til you’re done…
OK, I hope those books were as thought-provoking to you as they were to me! Now let’s take a look at the health of bees and beekeeping from ecological and economic perspectives.
It’s All About Economics
Again and again, I find that everything boils down to economics and finding the right niche. This applies to both honey bees and to the business of beekeeping—either thrives in its ideal niche, and either must either adapt or die if the parameters of the niche change. And boy howdy, how we have changed the parameters of both of our niches in recent years!
Practical application: In this real world, each species, and each business, strives to exploit a niche to which it is particularly well adapted. A change in any of the parameters that define a particular niche may affect the profitability and survival of that species or business. If that species or business is efficient and profitable in its particular niche, then it thrives; if not, if must either adapt or go extinct.
For the remainder of this article, I will view the situation of both bees and beekeepers through the lenses of ecology and economics, and the changes that have occurred in the parameters that define our niches.
Let’s Define Some Terms
Pollinators are in decline over much of the world, and have been for some time [8]. We beekeepers are mainly concerned with our favorite pollinator, the European honey bee, Apis mellifera, native to Europe and Africa, but now introduced worldwide. Unless I specify otherwise, henceforth I will be referring to this species.
It occurs to me that if pollinators have long been in decline worldwide, then that would imply that something has changed in their ecological niches (and that it started before the introductions of cell phones, neonics, or GMO’s). It also occurred to me that the niches occupied by beekeepers have changed substantially (mine sure has; indeed several times). I’ll try not to burden you with too many new terms:
Habitat—where the bee species lives (or could live). The bees in the U.S. are mongrel hybrids of various European or African races [9], each originally adapted to specific microhabitats in their home countries.
Ecological niche—a description of the bees’ “occupation” in its specific microhabitat, including all environmental parameters and interactions with other species.
Fundamental niche— the potential full range of environmental conditions and resources that the honey bee as a species could possibly occupy and use, without the limitations of predation, competition, or other factors.
Realized niche—the less-than-optimal niche that each subspecies of bee actually occupies; constrained by weather, resources, parasites, etc. In its home range, various subspecies of honey bee adapted to narrow realized niches occurring in the warm Mediterranean, the cold Alps, the British heathland, the Egyptian desert, the African savannah, etc. In each of those niches, the bees adapted to the seasonality of local nectar flows, the local plant toxins, temperature, predators, and parasite pressure. Conditions may not have been optimal, but each subspecies was economically successful at “making a living” within those parameters.
So let’s list the most important parameters of the fundamental niche of the European honey bee:
- It is a colonial species, existing as a superorganism with generally a single reproductive queen, supported by multiple patrilines of sterile workers, each exhibiting slightly differing genetics, behaviors, and resistance to parasites, toxins, and diseases (this within-hive diversity is extremely important, but often ignored by beekeepers).
- It is a generalist species, able to gather food resources from a wide variety of plants. As such, it is adapted to metabolizing a wide range of toxic plant alleleochemicals (and by extension, synthetic pesticides).
- It is primarily a pollinator; its diet normally consists solely of nectar and pollen, although those raw foodstuffs are processed into other products (honey, beebread, and jelly) for the actual consumption by the majority of the members of the superorganism.
- The bee cannot forage unless the ambient temperature is above roughly 55°F (12°C). This limiting factor constrains its range to those areas that have adequate bloom available when the temperature exceeds that value; any colony with hungry brood when daytime temperatures do not exceed 55°F will soon become stressed due to an inadequate supply of protein (this is a huge management tip).
- Unlike other insects, the European honey bee stores vast quantities of processed food for later consumption when resources are scarce.
- This allows the colony to do something that no other species of temperate insect can do—maintain an elevated body temperature, and rear brood, throughout the winter.
- In order to maintain that colonial body temperature, the European honey bee requires a protective insulated cavity within which to nest (Fig. 2).
- Somehow, the European honey bee evolved with remarkably few parasites—the only significant ones being Nosema apis, the bacteria causing the two foulbrood diseases, the fly Braula, rare infection by two opportunistic fungi in pollen (chalkbrood or stonebrood), and what were normally “economically unimportant” infections by any of several insect viruses.

Figure 2. These bees are at the top of the winter cluster at the interface between empty cells and sealed honey. Despite the air temperature being below freezing, the temperature of the bees beneath the surface was 67°F (19°C). This remarkable technique of using stored sugars from the previous summer as an energy source during winter allows the honey bee to overwinter as a populous colony, ready to exploit early spring pollen and nectar sources.
In summary, the honey bee requires a dry cavity, in which it maintains a tropical environment throughout the year, allowing it to exploit food sources any time that the temperature is above 55°F, it is adapted to metabolize a wide variety of plant toxins, it only requires a brief honeyflow to provide it with food stores for the remainder of the year, and it evolved under low parasite pressure.
Practical application: this last point is of huge import when we attempt to understand the biological changes that have occurred in European honey bee populations worldwide in the last decades. To wit, virus infections have become serious “emerging diseases” [10].
Add The Beekeeper
OK, now let’s add one more term:
Facilitation—Optimal conditions in the fundamental niche occur infrequently, if ever. The job of the beekeeper is to optimize the conditions for his bees as best he can, such as by supplemental feeding, medicating for parasites, protecting from predators, providing a larger nest cavity, or providing water or winter insulation. Such “facilitation” may allow bees to survive outside of their fundamental niche.
Practical application: I recently enjoyed a lunchtime conversation with a couple of professional beekeepers who moved their hives from almonds, to the tallow bloom in Texas, to the Dakotas for summer, and then to a mild area in California for wintering (their problem is having too many bees each spring). What they are doing is facilitating the optimal fundamental niche for their bees for the entire year (there are many other ways of doing so).
Practical application flip side: If a beekeeper is keeping colonies alive outside of their fundamental niche, such as in densely-packed apiaries, in areas of crop monoculture or high exposure to toxic chemicals, in flowerless forest or dry grassland, or by the chemical suppression of parasites, should that beekeeper falter in his constant facilitation, his bees may not be able to continue to survive without such help.
Limiting Factors
The realized niche of the honey bee is constrained by limiting factors, which may change from season to season. Common limiting factors for populations of honey bees are:
Climate—bees have very wide “tolerance limits” for cold, heat, rain, and length of seasons. But at the edges of their tolerance limits, colonies will be stressed, and may not be able to deal with other limiting factors.
Competition for food—in some areas there is such an abundance of nectar during the main flow that there is little competition (an important point when speaking with native pollinator advocates). The main competition for food resources occurs at other times of the year; assume that there is serious competition happening if robbing behavior is evident.
Suitability of available food—not all plants produce honey-bee-friendly nectar or pollen, especially outside of the honey bees’ native range. This is clearly evident in America and Australia, where some pollen sources are notably nutritionally inadequate for honey bees (think corn, blueberry, watermelon, pumpkin, some eucalypts). And in some areas or under dearth conditions, bees will unwittingly collect naturally toxic pollen or nectar. And of course, some human-applied pesticides make the available food unsuitable.
Competition for nest sites—without hollow trees or other natural cavities, honey bees cannot survive the winter in temperate climates (Fig. 3).

Figure 3. One of my colonies swarmed late this spring and built open-air combs in a nearby hawthorn tree. We noticed it when the leaves fell in early December, following a week of subfreezing temperatures and a foot of snow. You can’t see it, but there is still a cluster of live bees (which I hope to rescue when I’m done writing this article). The population had obviously grown large enough to build and completely cover all the combs, and could easily have survived the winter had it only found an appropriate cavity in which to build its nest.
Predation—Such as birds, bears, wasps, and ants. The main predator of bees, of course, are humans, who often rob too much honey from the hive, resulting in winter starvation.
Parasitism—Again, natural populations of European honey bees appear to historically have been minimally affected by parasites under normal conditions. We will return to how this has changed.
Transmission of parasites—This is very density dependent—the more colonies within flight range, and the more competition for resources, the greater the transmission of parasites. The swapping of combs by beekeepers also changes this dynamic.
Toxins—Natural plant allelochemicals, soil metals, industrial pollution, agrochemicals, and recently, a huge influx of beekeeper-applied miticides.
Bees have a wide range of tolerance for some limiting factors, and more narrow ranges for others. Usually, several factors interact (sometimes synergistically) to limit bee populations.
Practical application: there is often a single limiting factor that is the determinant for colony survival. A concept used in ecology is “Liebig’s law of the Minimum” (Fig. 4). A beekeeper can work hard all season long to do everything he can for his bees, but should he overlook any single critical limiting factor, Liebig’s Law may come into play, and he may lose his colonies.
Figure 4. An illustration of Liebig’s Law of the Minimum as it applies to the practice of beekeeping. Despite everything else that you do to fill the barrel, the most unfavorable limiting factor(s) (or some combination thereof) at any critical period of time will limit the bees’ (and your) success. Adapted from [[i]].
[i] Barrel illustration after Dobenecks, taken from Wikipedia http://en.wikipedia.org/wiki/Liebig%27s_law_of_the_minimum
Practical application: Each race of honey bee in Europe specialized by adapting to certain combinations of limiting factors, and thus gained a fitness advantage in its particular habitat. Since the arrival of varroa, which wiped out much of the feral population, the overall genetics of the U.S. bee population have likely shifted toward those propagated by commercial queen producers [12].
These “all-purpose” bee stocks are typically bred for color, temperament, and honey production, and maintained with a high degree of facilitation by the queen producer (feeding, treatments). There is no reason to expect those stocks to be well adapted for survival without constant facilitation. This is why I strongly support regional queen breeding for locally-adapted stock.
What Are The Limiting Factors For Honey Bees Today?
It would have been so simple had CCD actually been caused by cell phones! We could have banned the danged things, and wouldn’t have to listen to people walking around loudly and obliviously talking to themselves. But alas, it appears to be more complex than that.
So as a biologist, it occurs to me to go back before CCD, in fact, to go back even further in time, and ask the question, “Which factor(s) limited honey bee populations in Europe prior to modern management by humans”? And then we can work forward in time to see what’s changed since then. To be continued…
Footnotes and Citations
1 Directed by Jeremy Simmons (2009), and recommended for those who didn’t experience the horror of CCD personally. Unfortunately, I can’t suggest anywhere to purchase a copy of this well-done and heart wrenching film.
2 Pollinator Research.–The Committee is aware that pollinators are responsible for the production of one-third of the Nation’s food supply, but the number of managed honeybee colonies in the United States has dropped in half since 1940. Because of the importance of pollinators in the production of the Nation’s food supply and their impact on the stability of our agricultural economy, the Committee encourages [the Agricultural Research Service] to continue to dedicate resources to protecting the health of both honeybees and other native bees, including continued research into colony collapse disorder. http://www.gpo.gov/fdsys/pkg/CRPT-112srpt73/html/CRPT-112srpt73.htm
3 Symptoms of CCD:
1) In collapsed colonies
a) The complete absence of adult bees in colonies, with no or little build up of dead bees in the colonies or in front of those colonies.
b) The presence of capped brood in colonies.
c) The presence of food stores, both honey and bee bread
i) which is not immediately robbed by other bees
ii) when attacked by hive pests such as wax moth and small hive beetle, the attack is noticeably delayed.
2) In cases where the colony appear to be actively collapsing
a) An insufficient workforce to maintain the brood that is present
b) The workforce seems to be made up of young adult bees
c) The queen is present
d) The cluster is reluctant to consume provided feed, such as sugar syrup and protein supplement
From vanEngelsdorp, D, et al (2006, revised Jan 5, 2007) Investigations into the causes of sudden and alarming colony losses experienced by beekeepers in the fall of 2006. Preliminary Report: First Revision.
4 Williams, GR, et al (2010) Colony Collapse Disorder in context. Bioessays 32(10): 845–846. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3034041/
5 Janabi, F (2013) From anti-GMO to pro-science: ‘A Layman’s Guide to GMOs’. http://www.geneticliteracyproject.org/2013/12/03/from-anti-gmo-to-pro-science-a-laymans-guide-to-gmos/#.UqkDXdJ_dyI
6 Heinrich, B (2004) Bumblebee Economics. Harvard University Press. I highly recommend all of Heinrich’s books—he’s a brilliant scientist and an engaging writer whose passion it is to understand the details of how organisms survive in their niches.
7 Miksha, R (2004) Bad Beekeeping. Trafford. For a more data-based analysis, see:
Laate, EA (2013) Economics of Beekeeping in Alberta 2011. http://www1.agric.gov.ab.ca/$Department/deptdocs.nsf/all/agdex14472/$FILE/821-62.pdf
8 CSPNA &NRC (2007) Status of Pollinators in North America. https://download.nap.edu/login.php?record_id=11761&page=%2Fdownload.php%3Frecord_id%3D11761
9 The evolutionary origin of the European honey bee is currently under debate by scientists. See the following:
Han, F, et al (2012) From where did the Western honeybee (Apis mellifera) originate? Ecol. Evol. 2(8): 1949–1957. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3433997/
Kotthoff, U, et al (2013), Greater past disparity and diversity hints at ancient migrations of European honey bee lineages into Africa and Asia. Journal of Biogeography 40: 1832–1838. http://onlinelibrary.wiley.com/doi/10.1111/jbi.12151/pdf
10 Genersch, E & M Aubert (2010) Emerging and re-emerging viruses of the honey bee (Apis mellifera L.). Vet Res. 41(6): 54. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2883145/
11 Barrel illustration after Dobenecks, taken from Wikipedia http://en.wikipedia.org/wiki/Liebig%27s_law_of_the_minimum
12 Delaney, DA, et al (2009) Genetic characterization of commercial honey bee (Hymenoptera: Apidae) populations in the United States by using mitochondrial and microsatellite markers. Ann. Entomol. Soc. Am. 102(4): 666-673.
First published in: American Bee Journal January 2013
Sick Bees Part 18F1
Colony Collapse Revisited
Pesticide Kills
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ in January 2013
Pesticides
A Bit of History
Nailing Down the Guilty Party
Keep ’em Honest!
Pesticides and CCD
Making the Link
Acknowledgements
References
As long as I’ve been keeping bees, one of our worst fears has been that we might suffer a serious pesticide kill. Pesticides (especially insecticides) have always been, and will continue to be, a problem for bees and beekeepers.
Jim Doan of New York hadn’t experienced a serious pesticide kill in 25 years of keeping bees in corn/soy/alfalfa farmland. But when he approached one of his yards last spring, he smelled the stench of dead bees. What he saw made him sick to the stomach—piles of dead and rotting bees in front of every hive!
Jim related to me that he called his state’s Department of Conservation to investigate the kill, to no avail. So he contacted his State Apiarist, who sent out an inspector a couple of days later to take samples, which then languished in a refrigerator until at Jim’s request they were sent to Dr. Maryann Frazier, who in turn sent them to the USDA lab for analysis.
Although pesticide residues were found, no investigation was done. No applicator was reprimanded, and no fine imposed. And Jim’s losses weren’t covered by either his insurance policy or by ELAP [1].
Jim’s disaster was hardly an isolated case. I’ve spoken with a number of beekeepers who have suffered recent pesticide kills. Dave Shenefield’s bees were working white clover in Indiana at corn planting time. A farmer drilled treated corn seed directly into a field of flowering clover without first burning the weeds off with herbicide. The planting dust fell directly onto the blossoms being worked by the bees, poisoning his colonies as the foragers returned covered with toxic dust.
Darren Cox’s bees in Utah get hit regularly by applications of pyrethroids or carbamates onto flowering alfalfa hay. These applications done are despite label restrictions that clearly state:
“This product is highly toxic to bees exposed to direct treatment or residues on blooming crops or weeds. Do not apply this product or allow it to drift to blooming crops or weeds if bees are visiting the treatment area.”
Darren related to me a scenario: an aerial applicator, under a contract arranged perhaps two weeks earlier, loads up with insecticide, and flies 50 miles to treat the field. But when he gets there, he’s surprised to see that the alfalfa is purple with bloom. What’s he to do—turn around and unload, or just go ahead and spray anyway, knowing that such an action would be in violation of the label. But this is Utah, where the local primacy partner tends to turn its head to pesticide violations (Fig.1). You can guess the rest–such unnecessary and preventable bee kills frustrate Darren to no end!

Figure 1. A typical insecticide kill in Utah. Simple timely communication between the grower and the applicator as to the stage of bloom could prevent many such kills, as the applicator could then make more appropriate product application choices. Photo courtesy Jared Taylor.
I could go on and on, beekeeper after beekeeper. What I hear is that some states are better at others at enforcing pesticide regulations—it’s tough to be a beekeeper in those states that aren’t doing their job! To make things worse, beekeepers are often justifiably hesitant to pursue investigation, since in a number of states, complaining beekeepers have been fined for having illegal miticide residues in their hives! And if a beekeeper raises too much of a stink he could become persona non grata to the local landowners and lose his locations.
Farmers and applicators could often easily prevent bee kills by simply making sure that they spray before or after a crop comes into bloom, or by spraying after dusk with a product having a short residual toxicity, or by using a less bee-toxic product that is labeled for application during bloom. Such practices would eliminate a large proportion of bee kills, yet some farmers and applicators just don’t give a damn, and worst of all, get away with illegal applications (scofflaw applicators may consider any fines levied for pesticide misapplication as a minor business cost)!
What bothers beekeepers most is the unfairness of it. Ranchers (even of alpacas, reindeer, or emus) receive government benefits for livestock losses due to fire or severe weather [2], and beekeepers may be eligible for benefits for colony losses if they jump through the hoops of ELAP [3]. But neither of those programs cover losses due to pesticide application–either legal or in violation of the law.
Think about it–if someone poisoned a herd of cattle with pesticide overspray, it would make the news! You could damn well bet that the incident would be investigated and the applicator fined, and the cattleman would sue for damages via civil action. But this is generally not the case if your livestock are honey bees. Few damaged beekeepers receive any compensation at all for their losses.
Now I don’t want to give the impression that the pesticide situation is dire for all beekeepers. As I pointed out in a previous article [4], many beekeepers in agricultural areas have little or no problem with pesticides. And many commercial beekeepers simply shrug off the occasional bee kill as a cost of getting good locations in agricultural areas. However, in some areas of intensive agriculture, those commercial beekeepers who provide the bulk of pollination services tell me that pesticide issues are their major problem.
A Bit of History
In order to understand the run up to our current situation, it is helpful to read the engaging “Report on the Beekeeper Indemnity Payment Program” (which was in effect from 1967-1980) [5]. I’ll share a few excerpts:
During the mid-1940’s, [pesticide] damage subsided as farmers shifted from the use of arsenicals to DDT which is less toxic to bees. However, by the late 1960’s, use of DDT was decreased sharply because of insect tolerance to the poison. Finally, use of DDT and other chlorinated hydrocarbons was banned because of environmental concerns. In most cases, the highly toxic [organo] phosphates and carbamates were used in place of the banned sprays. This increased the problem of bee loss to the point of disaster for many beekeepers…
Partial colony losses are not always easy to detect…pesticides may weaken colonies to such a point that they do not survive the winter. This type of loss is often ascribed to winterkill rather than pesticides. Further, this loss may be extended to the replacement bees placed in contaminated equipment the next season. Often, not all losses are discovered soon enough after the chemical application to determine the exact cause of death.
Investigatory clue: these records of the field experiences of beekeepers prior to varroa are important to keep in mind, notably that there were “sublethal effects” from the pesticides that caused later winter mortality. I hear the exact same complaints from beekeepers in agricultural areas today. Clearly, varroa and beekeeper-applied miticides have added to the stress upon bee colonies, but elevated winter mortality due to pesticide exposure was the norm prior to the introduction of varroa.
Colony losses due to pesticides were severe in several states during the 1960’s. There was a “sharp decline in pesticide losses” in California during the early ‘70’s due to the state imposing “strict control of spray application”—only 54,000 colonies were killed in 1974, compared to 89,000 in 1970 (an improvement, but hardly cause for celebration). But then in the mid 1970’s, encapsulated insecticides (Penncap-M) were brought to market, again causing devastating losses when foragers dusted with the time-release particles returned to the hive and stored them in the beebread.
During June 1976, selected beekeepers in California and Washington were contacted to discuss the pesticide situation…Beekeepers in Washington report that there are no safe locations for bee yards. One beekeeper said, “No matter where I place my bees in the Yakima Valley, they will be sprayed at least once within ten days.” A beekeeper in the San Joaquin Valley of California described his efforts to protect his apiaries as “playing musical chairs with 40 loads of bees….” Several beekeepers said that even if they did move their colonies to another location, it could be sprayed the next day.
Practical application: I hear exactly the same words today from commercial operators. We have made great progress with pesticides since the 1960’s, but still not enough!
Beekeepers in Arizona, California, and Washington accounted for a large proportion of claims because they lacked access to “safe” forage areas (these were the early days of using forklifts in bee operations, and moving bees was hard work). It was not unusual for large beekeepers to suffer serious pesticide damage to half their hives each year, and they would likely have been unable to stay in business without governmental help (Fig. 2).
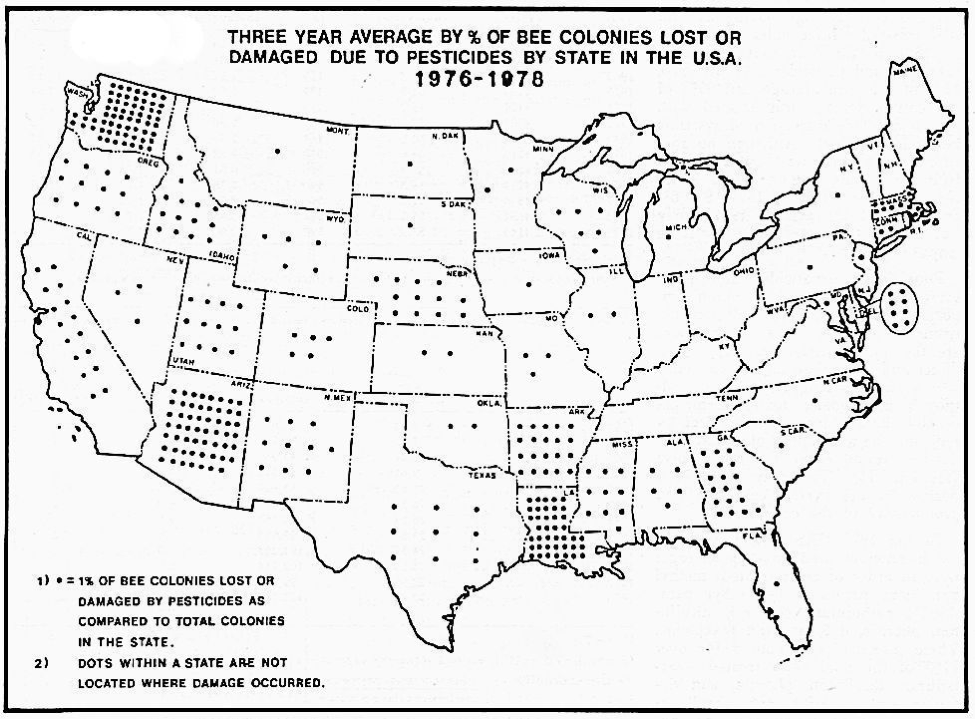
Figure 2. Back when the Agricultural Stabilization and Conservation Service kept records of reported bee kills for indemnification purposes (not all kills were reported), it was easy to see in which states pesticide applications were a serious problem. In recent years bee kills have not been tracked by any agency. Map from Erickson & Erickson 1983 [[i]].
[i] Erickson, EH, and BJ Erickson (1983) Honey bees and pesticides. ABJ 123(10): 724-730.
For nearly a decade, the Indemnity Program compensated beekeepers for pesticide losses. Those in only eight states filed the bulk of claims. As today, a small percentage of commercial beekeepers control the vast majority of colonies, and provide most pollination services. Well less than 1% of beekeepers in the country filed claims in any one year. By contrast, over 90% of the Arizona beekeepers in the program filed claims— not surprising due to the frequent spraying of the vast acreage of cotton suffering from a serious infestation by pink bollworm in the mid 1960’s [7], and the lack of alternative non-agricultural forage in that dry state.
The largest payment to a single beekeeper (name and state not specified) was $225,400 in 1972 (that would be $1 million in today’s dollars), and he filed for $228,000 two years later. You can imagine how this might not have set well with some budget-conscious congressmen!
And of course some crafty beekeepers learned to work the system:
On the other hand, some commercial beekeepers contend the indemnity payments have permitted, and in some cases encouraged, the survival of marginal beekeeping operations. The “marginal manager,” in this context, was characterized as any beekeeper who had become dependent upon indemnity payments as a source of income.
Those alleged “marginal beekeepers” reportedly left their hives in areas that they knew would be sprayed, and managed their colonies only enough to keep them barely alive so as to be able to collect more payments the next year (or kept collecting payments on deadouts). These fraudulent practices also did not play well to the program overseers.
The study also looked at the profitability of beekeeping; I found one of the tables to be of particular interest (Fig. 3):
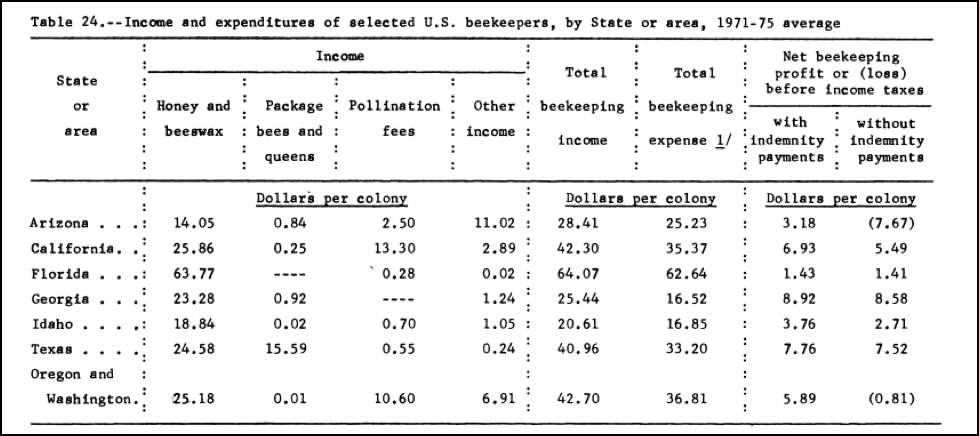
Figure 3. You can roughly adjust these figures into today’s dollars by multiplying them by five. What surprises me is that despite it being painfully costly to maintain colonies today in California (the annual expense being about $190) [[i]], the profit margin is substantially higher now than it was back then–not because of honey (since honey prices have only kept pace with inflation [[ii]]), but rather due to much higher pollination rates in almonds. Also of interest is that in those days beekeepers spent next to nothing on feeding syrup, and pollen supplement isn’t even mentioned!
[i] Mussen, E (2009) How much does it cost to keep commercial honey bee colonies going in California? http://projectapism.org/content/view/83/27/
[ii] http://www.nass.usda.gov/Statistics_by_State/California/Historical_Data/Bees.pdf
It is instructive that the analysts were aware of the cost to the beekeeper of pesticides:
This analysis shows that beekeeping income is affected most by severely damaged and destroyed colonies. Severely damaged colonies may require 6-8 weeks to recover colony strength. If the damage occurs during a major honey flow, the field force will be greatly reduced and honey yields could be lowered 60 percent or more. Severe damage in late summer may weaken a colony preparing for winter and increase the chances for significant winter kill…Beekeepers estimate it takes about one year for a destroyed colony to regain its income earning potential.
The authors conclude that without the indemnity payments, “farmers seeking pollination services would have to pay substantially higher rental fees to obtain bees.” Congress decided to pass that cost onto the farmers anyway, and terminated the indemnity program in 1980 (leaving some beekeepers with still-unpaid IOU’s). Today the almond growers bear the brunt of those higher rental fees; the huge number of colonies produced to meet the demand for high-paying almond pollination ensures that there are plenty of strong hives available for other crops afterwards.
Colonies generally come out of almonds in better shape then when they went in. This is not true for a number of other crops. The combination of poor forage and pesticides in several crops can weaken colonies to the extent that they don’t survive the season.
Allow me to close with some prescient conclusions from the report:
Unless Federal and State governments act ot regulate and caution applicators of toxic pesticides, colony damage will continue to be a major problem for beekeepers. However, most government officials emphasize that farmers and spray applicators are already confronted with enough regulations…the current development of stronger and longer-lasting pesticides…is creating an environment entirely unsuitable for honey bees in many parts of the U.S. These areas will find it harder to maintain the present level of bee population regardless of an Indemnity Program or higher honey and pollination prices.
Remember that the above words were written prior to the invasion of the tracheal mite, the varroa mite, Nosema ceranae, or the Small Hive Beetle—beekeeping hasn’t gotten any easier since their arrival!
Practical application: beekeeping in agricultural settings has always been a tough way to make a living. Fortunately, many beekeepers tell me that things have gotten better in their regions. But in some areas of intensive pesticide application, it’s hard to keep a hive alive from one year to the next.
Nailing Down the Guilty Party
This spring my bee operation suffered from a case of Sudden Forklift Collapse (Figure 4).

Figure 4. Early this spring I suffered from a case of Sudden Forklift Collapse. This was no “sublethal effect” and did not go unnoticed! In a forklift kill like this, it didn’t take Sherlock Holmes to determine that the cause of death was due to a falling oak tree. If only the causes of pesticide kills were so easy to pin down!
In my case of Sudden Forklift Collapse, the cause was evident. Such is often not the case with pesticide kills. You may not even see any dead bees if the field force is poisoned in the field and never makes it back to the hive. Perhaps (as in the case of planting dust) you only see a handful of young bees and drones dying at the landing board. Or maybe the brood turns spotty. If the pesticide disorients the foragers, you may wonder why you didn’t get the normal honey crop. Or maybe there is some sublethal effect from which the colony simply “slows down” for a few months, or doesn’t make it through the winter.
In any of those cases, it may be difficult, if not impossible, to nail down the culprit. You don’t know where your bees were foraging, and any pesticide application within a 3-mile radius is suspect. You may not immediately recognize that there was a pesticide problem at all, so any residues could be degraded or washed off by rain by the time you think to have the dead bees or beebread tested. And even if you happen to visit the yard immediately after the kill, good luck in getting an understaffed and untrained state or county agency to quickly come out and properly collect and freeze a good fresh sample. And even then the analytical tests cost so darned much!
Action item: aggrieved beekeepers often have VERY STRONG FEELINGS! However, in order to change pesticide regulations, the EPA needs incontrovertible evidence that a certain pesticide used according to label restrictions caused adverse effects to honey bees. We need any and all beekeepers who suffer from substantial pesticide kills to file an “incident report.” Such a report is most effective if it contains a photographic record, documentation that rules out other plausible causes for the dead bees (e.g., tracheal mites or starvation due to unusual weather or forage conditions), and chemical analysis of samples of bees and beebread, properly taken by a state agent. If your local primacy partner is unable or unwilling to help, you may report directly to http://www.npic.orst.edu/eco or beekill@epa.gov.
One would think that solving Jim Doan’s kill would have been straightforward, since there were fresh piles of dead bees in front of the hives. He hadn’t previously experienced serious kills in those yards, so something different had happened. There was no apparent change in plantings this year, but with commodity prices at an all-time high, a farmer might have felt that it was worthwhile to apply more or different insecticides as precautionary “risk management.” Surely it would be easy to find incriminatingly-high levels of the offending pesticide in the dead bees or combs.
According to Jim, due to unfamiliarity with the investigation of pesticide kills, the state inspector collected less than an optimal amount of bees for pesticide analysis. Two samples were later sent off to the USDA lab (the cost of analysis was split between Jim and Project Apism)—results below (Fig. 5).

Figure 5. Analysis report of the two samples from Jim Doan’s spring bee kill (column headings added).
Figure 5. Analysis report of the two samples from Jim Doan’s spring bee kill (column headings added).OK, so now Jim had a report. But what did it tell him? As for the dead bees, the 1.6 ppb* of clothianidin insecticide is far too low to have caused bee mortality (1.6 ppb = 0.16 ng/bee; the LD50 for clothianidin lies in the range of 22-44 ng/bee).
* To help with the math, LD50 = median lethal dose; 1 ppb = 1 part per billion = 1 μg/kg = 1 ng/g; μg = microgram (one millionth); ng = nanogram (one billionth); a bee weighs about a tenth of a gram, so for every 10 ppb of residues in a sample of dead bees, any bee on average would contain 1 ng/bee .
So how about the high dose of Captan fungicide? As best I can tell from the literature, “Studies on the honeybee using technical Captan fungicide indicate that the LD50 is greater than 10 μg a.i./bee, and that there is 9.8% mortality at 215 μg a.i./bee.” So let’s do the math: 1290 ppb = 129 ng/bee, or 0.129 μg/bee—so again, it would be hard to make a case that this chemical was responsible for the obvious pile of dead bees.
Maybe the analysis of the pollen sample from the comb might help. I have no idea as to how it was taken, which can make a huge difference (Fig. 6).

Figure 6. These are plugs of beebread that I pulled from a brood frame. Note the layering of the different species of pollen. If a colony suffers from a pesticide kill, any traces of the responsible pesticide residue may only be in the topmost layer of pollen. If the state agent who takes the beebread sample scoops all the way to the midrib, he may dilute the offending pesticide by a factor of 10 or more.
The one pollen sample from the one comb from one colony (get my point?) in Jim’s affected apiary contained 399 ppb of the organophosphate insecticide Phosmet. The contact LD50 for this compound is listed as 0.0001 mg per bee (= 0.1 μg/bee = 100 ng/bee). Surprisingly, there doesn’t appear to be any published oral LD50 for Phosmet to honey bees! By my math, the concentration of Phosmet in Jim’s pollen sample would not be expected to have killed his bees either, although since it is a violation of the label to spray the insecticide on flowering crops, one is left wondering how it appeared in the pollen.
So this is how it can be for a beekeeper and his innocent bees—the suddenly-appearing piles of rotting corpses in front of every one of his hives certainly suggest that his bees were killed by a pesticide application. Unfortunately, due to a lackluster investigation by the primacy partner, and lack of implicating chemical evidence, Jim will never know what or who was responsible for the kill, nor be compensated for his losses, if justified. And he has no idea whether the same thing will happen again next season!
To make matters worse, Jim’s bees apparently got hit again in July, resulting in piles of greasy-looking dead and twitching dying bees in front of the entrances. And as I write these words in November, Jim sent me yet another photo of hundreds of freshly-dead bees once again in front of the hives (despite him confirming low levels of varroa and nosema). Jim is now a justifiably frustrated and angry beekeeper–not only did he suffer considerable financial loss (not to mention the ugly death of his beloved bees), but no one learned anything from the experience! The unwitting farmer(s) have no idea whether their pesticide applications caused the problem, Jim’s state agencies aren’t making any particular effort to prevent the same thing from happening again next year, and EPA didn’t receive any useful adverse effects report. Yes, frustrating!
It is disturbing for me to present these facts. Our managed honey bees function as a conspicuous and charismatic indicator species for the effects of pesticides upon “non target organisms.” Yet some agricultural areas are a “no bees land” due to either inadequate label restrictions or flagrant violation of those restrictions. And keep in mind that the honey bee colony has the capacity to absorb pesticide kills that would exterminate solitary pollinators, such as native bees, butterflies, and beneficial insects.
Practical application: if honey bee colonies are being killed, we can safely assume that the situation is even worse for more sensitive species!
Keep ‘em Honest!
Let me share another quote from the Indemnity Report:
The Beekeeper Indemnity Program itself discourages civil court action…Greater use of the civil court system by beekeepers to seek compensation for pesticide losses could reduce applicator negligence.
There you have it! The sad truth is that it will take the push of lawsuits to ensure that our pesticide laws are actually enforced. Accordingly I’ve studied the judgments for some beekeeper lawsuits. Be forewarned that a successful lawsuit requires unimpeachable evidence and impeccable argumentation—so one should not enter into an expensive lawsuit lightly!
The AHPA has started a legal defense fund to pursue test cases against egregious violations of pesticide law, with the hope of setting legal precedent, as did Jeff Anderson’s successful lawsuit against the state of Minnesota in 2005 [10]. I’m hesitant to step into politics, but I feel that this is probably a good course of action that could help the cause of advancing pesticide regulation. We beekeepers must tread carefully here to avoid pissing off the farmers who allow us to place bees upon their land. In truth, I’d like to see Xerces or some other environmental groups filing such lawsuits, so that they, rather than beekeepers, would take the heat. However, action is preferable to inaction.
Action item: you may join me in contributing to the National Pollinator Defense Fund at http://pollinatordefense.org/site/?page_id=11
I wish that I could present a simple solution to this problem, but there isn’t one—especially since the U.S. is currently locked into the high-input large-scale monoculture agribusiness model. The good news is that EPA is on the side of the beekeepers and the environment [11], and that things are clearly getting better—the worst pesticides are being phased out, new “reduced risk” pesticides and “biological” are put on the EPA fast track in order to get them into the market, plus a new generation of “smart” robotic application systems are being developed. There has never been more public awareness of the plight of the honey bee, and beekeepers are awkwardly basking in the spotlight of being considered as environmental stewards. The bad news is that the process of reducing the damage by pesticides to non target species is hampered by, among other things, ignorance (and lack of enough good scientific data), politics, property rights, consumer demand, and Money (intentionally spelled with a capital M).
OK, that’s enough griping for now–let’s get back to an investigation into any connections between pesticides CCD.
Pesticides and CCD
Biological plausibility: pesticides can weaken the colony by killing or otherwise affecting the foragers, reducing adult bee longevity, having adverse effects upon the queen, brood, or nurse bees, or by affecting bee behavior. In addition, they could react with other toxins, beekeeper-applied miticides, or suppress the bee immune response to pathogens. Any of the aforementioned could conceivably result in colony dwindling, mortality, or collapse.
Residues in the Combs
Let’s narrow down our focus. CCD by definition is not the result of the sorts of acute pesticide kills detailed above. So what we are interested in is colony mortality or morbidity due to sublethal effects that hadn’t already killed bees outright! In the case of winter mortality, since few pesticides are applied at that time of year, and since colonies normally purge any remaining field bees during the “fall turnover” [12], we’d expect any contribution by pesticides to be from residues in the combs, where they should be detectable by analysis.
Making the Link
One would think that it would be a simple matter to make the connection between pesticide residues and winter mortality—simply analyze pollen and beeswax samples from the combs, and determine whether there is a correlation between residues of specific pesticides and colony mortality.
The above sounds so straightforward and easy, but in actuality this is where it gets complicated. My point of going into detail on the analysis of Jim Doan’s apparently obvious bee kill was that if it’s that hard to figure out exactly what caused an acute pesticide kill, imagine how difficult it would be to definitively link colony mortality to any sublethal effects from a specific pesticide!
In fact, I took artistic license in greatly simplifying Jim’s story. In doing my usual fact checking, I found out that the actuality was complicated by personalities, politics, weather (Fig. 7), and a history of indemnity payments. To add further confusion, another beekeeper on the same farm did not observe any dead bees in front of his hives (but did notice that his nucs on that farm did not build up as well as those at other nearby locations).
Figure 7. Western New York experienced extraordinarily warm weather (followed by cold) in May. I find that such weather anomalies can result in piles of dead bees in front of hives due to short-term starvation events. Weather graph from www.weatherunderground.com.
However, I’m appreciative of Jim for sharing his observations and analysis report, and feel that it was a good example of the problems that researchers and regulators encounter as they try to figure out exactly how pesticides are affecting colony health.
These complicating factors may be why no scientific study has yet been able to firmly link colony mortality to pesticides. Here are the conclusions of all monitoring and analytical studies that I’ve seen to date:
- Germany: “As expected, the results show that pollen [from 210 hives sampled over 3 years] is contaminated with a plethora of chemical substances originating from the agricultural practice of using pesticides but also from the apicultural necessity of using acaricides… Accordingly, no relation between contamination of pollen and colony development or winter losses could be demonstrated in the course of the project although special emphasis was put into this aspect” [13].
- France: “Several cases of mortality of honey bee colonies (varying from 38 to 100%) were observed in France during the winter of 2005-6. In order to explain the causes of these mortalities, a case control study was conducted on a limited area, together with a larger survey in 18 other apiaries located in 13 sites over the entire country…No pesticide residues of agricultural origin were found in the samples of beebread, beeswax, honey and dead honey bees, with the exception of imidacloprid…found in one apiary [and] not considered to be able to cause honey bee acute mortality” [14].
- France: “A 3-yr field survey was carried out in France, from 2002 to 2005, to study honey bee … colony health in relation to pesticide residues found in the colonies… No statistical relationship was found between colony mortality and pesticide residues” [15].
- Italy: “The data obtained from the winter 2009-2010 inspections were used as the basis for chemical analyses on bee and wax samples, to test for residues of organophosphate, organochlorurate, carbamate and neonicotinoid pesticides, but no significant presence of these substances was detected” [16].
- Spain: “The present data [beebread samples from 12 apiaries] are in agreement with studies showing no negative effects of seed-treated crops. Some pesticide residues were found here, in particular several varroacides and insecticides, but no significant differences were observed between the different sunflower crop samples and those from the sites of wild vegetation. This fact not only implies environmental contamination but also supports the theory that, most of the time, inadequate [read that “unapproved”] treatments are the main source of residues that might weaken bee colonies and make them more sensitive to other factors” [17].
- Spain: “This study was set out to evaluate the pesticide residues in stored pollen from honey bee colonies and their possible impact on honey bee losses in Spain. In total, 1,021 professional apiaries were randomly selected… A direct relation between pesticide residues found in stored pollen samples and colony losses was not evident accordingly to the obtained results” [18].
- Europe (thorough review): “Currently there is no clear evidence from field based studies that exposure of colonies to pesticides results in increased susceptibility to disease or that there is a link between colony loss due to disease and pesticide residues in monitoring studies” [19].
- USA (CCD Descriptive Study): “This study found no evidence that the presence or amount of any individual pesticide occurred more frequently or abundantly in affected apiaries or colonies” [20].
- USA (2012 CCD Progress Report): “When pesticides are viewed in aggregate on a national scale, residues of pyrethroids …pose a threefold greater hazard to bee colonies than neonicotinoids, based on mean and frequency of detection in pollen samples and relative acute toxicity. The synthetic pyrethroid detected in the highest quantity and frequency in honey bee colonies that is used by beekeepers to control Varroa mite is tau fluvalinate” [21].
- USA (Stationary Hive Project) : “We did not find any relationship with any of our measures of pesticide contamination and colony loss rate at the apiary level for either 2009 or 2010” [22].
OK, I’m as puzzled as you are! It defies both common sense and a long history of beekeeper experience that researchers haven’t yet nailed down any link between pesticide residues in the combs and colony mortality! The above were not industry-funded studies, and several of the researchers started with a strong anti-pesticide bias (nearly all researchers suspect that pesticides are involved to some extent). And I’m certainly not about to tell you that pesticides/miticides and winter mortality are unrelated–it’s just, like I said, complicated.
I found that in order to begin to understand the effects of manmade pesticides upon bee health that I first needed to back up and examine some of the complex biology involved in natural bee/plant/toxin interactions. We’ll start in on that next month…
Acknowledgments
I’d like to thank the editor of this journal, Joe Graham, for giving me the latitude, support, and encouragement to write this series of articles. And a special thanks to Dianne Behnke of the publishing department for digging up and scanning archived issues of ABJ for my research.
References
[1] http://www.fsa.usda.gov/Internet/FSA_File/elap_honeybee_11.pdf
[2] http://www.fsa.usda.gov/Internet/FSA_File/lip2011_158c020211.pdf
[3] http://www.fsa.usda.gov/Internet/FSA_File/elap_honeybee_11.pdf
[4] https://scientificbeekeeping.com/the-extinction-of-the-honey-bee/
[5] ERS (1976) Report on the beekeeper indemnity payment program. http://babel.hathitrust.org/cgi/pt?id=coo.31924001799307;seq=8;view=1up
[6] Erickson, EH, and BJ Erickson (1983) Honey bees and pesticides. ABJ 123(10): 724-730.
[7] http://naldc.nal.usda.gov/download/48077/PDF
[8] Mussen, E (2009) How much does it cost to keep commercial honey bee colonies going in California? http://projectapism.org/content/view/83/27/
[9] http://www.nass.usda.gov/Statistics_by_State/California/Historical_Data/Bees.pdf
[10] Anderson v. State Department of Natural Resources Minnesotahttp://www.animallaw.info/cases/causmn693nw2d181.htm
[11] http://www.epa.gov/opp00001/ecosystem/pollinator/then-now.html
[12] Mattila HR, Otis GW (2007) Dwindling pollen resources trigger the transition to broodless populations of long lived honeybee each autumn. Ecol Entomol 32:496–505.
[13] Genersch E, et al (2010) The German bee monitoring project, a long term study to understand periodically high winter losses of honey bee colonies. Apidologie 41: 332-352.
[14] Chauzat MP, et al (2010) A case control study and a survey on mortalities of honey bee colonies (Apis mellifera) in France during the winter of 2005-6. Journal of Apicultural Research 49: 40-51.
[15] Chauzat MP, et al (2009) Influence of pesticide residues on honey bee (Hymenoptera, Apidae) colony health in France. Environmental Entomology 38: 514-523.
[16] Mutinelli, F, and C Porrini (2010) Report based on results obtained from the second year (2010) activity of the APENET project. http://ebookbrowse.com/apenet-2010-report-en-6-11-pdf-d189566755
[17] Bernal J, et al (2011) An exposure study to assess the potential impact of fipronil in treated sunflower seeds on honey bee colony losses in Spain. Pest Management Science 67: 1320-1331.
[18] Bernal J, et al (2010). overview of pesticide residues in stored pollen and their potential effect on bee colony (Apis mellifera) losses in Spain. Journal of Economic Entomology 103: 1964-1971.
[19] Thompson, HM (2012) Interaction between pesticides and other factors in effects on bees. http://www.efsa.europa.eu/en/supporting/doc/340e.pdf
[20] vanEngelsdorp D, et al. (2009) Colony Collapse Disorder: A Descriptive Study. PLoS ONE 4(8): e6481.
[21] http://www.ars.usda.gov/is/br/ccd/ccdprogressreport2012.pdf
[22] Drummond, F, et al (2012) The first two years of the stationary hive project: Abiotic site effects. http://www.extension.org/pages/63773/the-first-two-years-of-the-stationary-hive-project:-abiotic-site-effects



