



Understanding Colony Buildup and Decline: Part 9a – The Regulation of Bee Longevity
Understanding Colony Buildup and Decline: Part 9a
The Regulation Of Bee Longevity
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ in January 2016
CONTENTS
The Business Model Of The European Honey Bee
The Phases Of Colony Life Cycle
It’s All About Most Efficient Use Of Resources
Reversing The Question
The Regulation Of Aging
The Honey Bee As A Model For Gerontological Research
Bee Female Phenotypes
The Two Paths
Next
Acknowledgements
Notes And Citations
During the peak of the honey flow, the colony shifts from its primary springtime goals of broodrearing and reproduction, to honey storage and preparation for surviving the winter. In areas with a typical late-summer dearth of forage, this means reducing the colony population in order to minimize the number of mouths to feed.
The Business Model Of The European Honey Bee
As a biologist, I’m curious to learn the reasons that the bee colony does what it does at each time point of the year. As a beekeeper, the better I understand those reasons, the more likely I am to be able to work with my bees, rather than against them. There are reasons why the European honey bee is such a successful insect. And those reasons are the hard-wired behaviors honed by thousands of generations of natural selection by an unforgiving Nature.
To understand the European honey bee (EHB), we need to fully grasp the essence of its “business model”—which is to be the most efficient exploiter of the diverse floral bloom in cold-winter climates (especially that in early spring) (the African races easily outcompete the EHB in warm-winter climates). The EHB does this by having a fully-developed forager population at the ready early in spring, so as to be able to fully exploit the first flushes of spring bloom, thus giving it a jump on its competitors. By doing so, the colony can then reproduce (swarm) early enough for both the parent and daughter colonies to establish or restock their nests prior to the end of bloom.
The Phases Of Colony Life Cycle
Let’s consider the annual life cycles of social insects, which can be divided into phases [1]. I’ll take a bit of liberty here and divide the honey bee business model into four phases:
- The growth phase: the establishment of the broodnest, followed by rapid linear growth during favorable forage abundance (this is similar for either an overwintered colony or for a swarm).
- The reproductive phase: once a population size threshold has been reached (and conditions being favorable), the issuance of one or more reproductive swarms [2].
- The food storage phase: the storage, during the peak of bloom, of enough energy and protein reserves (in the form of honey and beebread) to last until forage becomes available again.
- Conservation mode: once forage becomes scarce, reduce the population and hunker down to maintain the queen and defend those stores until favorable conditions resume.
Maintaining a large population of sterile workers would no longer contribute to the business plan. In fact, once the colony has filled the nest with honey, any excess workers constitute nothing more than a bunch of hungry mouths to feed, no longer contributing to the colony’s goal—to get the queen through the winter.
This article comes as the culmination of my trying to understand how, and why, the colony regulates worker longevity. I chose to insert it into my series at this time, since this is when the colony makes a major shift.
It’s All About Most Efficient Use Of Resources
Survival of the honey bee colony in nature is all about efficiency in a highly competitive business arena. The efficiency of the superorganism is dependent upon the return on investment that it gets from each of its members.
Practical application: I suggest that for any of you considering making a living at keeping bees, to get a damn good grasp of the concept of “return on investment.”
Bees can be incredibly efficient at harvesting and storing honey, with every individual forager calculating the greatest return on investment for each trip [3]. Szabo [4] determined that during the Alberta honeyflow, colonies stored 24 times as much honey as the total body weight of the bees in the hive. Since only about half the bees in a hive are directly involved in nectar collection and processing, that’s about a 50 to 1 ratio of honey storage to worker body weight—pretty impressive!
Yes, efficiency in nectar collection and storage is critical, but colony population, on the other hand, is largely limited by the amount of incoming protein obtained from pollen. Protein is the most precious commodity in bee life, and the colony is frugal as heck with it. So long as pollen is abundant, the colony will remain in the growth phase, keeping the option of reproduction open–swarming as it approaches population top out, or if the broodnest plugs out with sealed brood (indicating that the colony has filled its cavity).
But when pollen becomes scarce, the colony responds immediately. How the information concerning a shortage of protein is communicated to all the members of the colony is both simple and elegant. As Seeley [5] nicely explains, “information flows between [the colony’s] members via mechanisms that are remarkably diverse and often curiously subtle.” The key communicators in this case are the nurse bees, via the exchange of the” currency” of protein—jelly. Since every bee in the hive other than nurses (and newly-emerged drones) depends upon begging jelly from nurses to fulfil their daily requirement for protein, the nurses can keep the queen, the foragers, and every other member of the hive up to date as to the status of the colony’s protein abundance.
When the nurses tighten up on the circulation of jelly, every bee in the hive knows that it’s time to conserve protein (and possibly let the foragers know to shift to pollen collection). As the protein deficit grows, drone brood is cannibalized, and then the colony summarily dismisses its adult drones [6]. At this point in time, a critical thing happens. The nurse bees start rearing the last generation of brood before the food runs out. That generation will become long-lived (diutinus) bees that “hold the fort” and protect the queen until pollen again becomes available. This typically occurs in fall after the first killing frost, but can also occur in arid regions during summer pollen dearth–in other words, at any time that no broodrearing (recruitment) is taking place [7].
Practical application: this is why good pollen nutrition is critical in the fall. Colonies that go into conservation mode without adequate protein reserves are unable to adequately deal with the stresses of wintering, nor can they sustain midwinter broodrearing.
Reversing The Question
When I present Lloyd Harris’ data (Fig. 1) showing that diutinus worker bees during winter may live for over 200 days, people often ask, how can a bee live for so long? But to me, the real question we should ask is: Why do workers normally live such short lives (averaging only around 35 days during spring and summer)?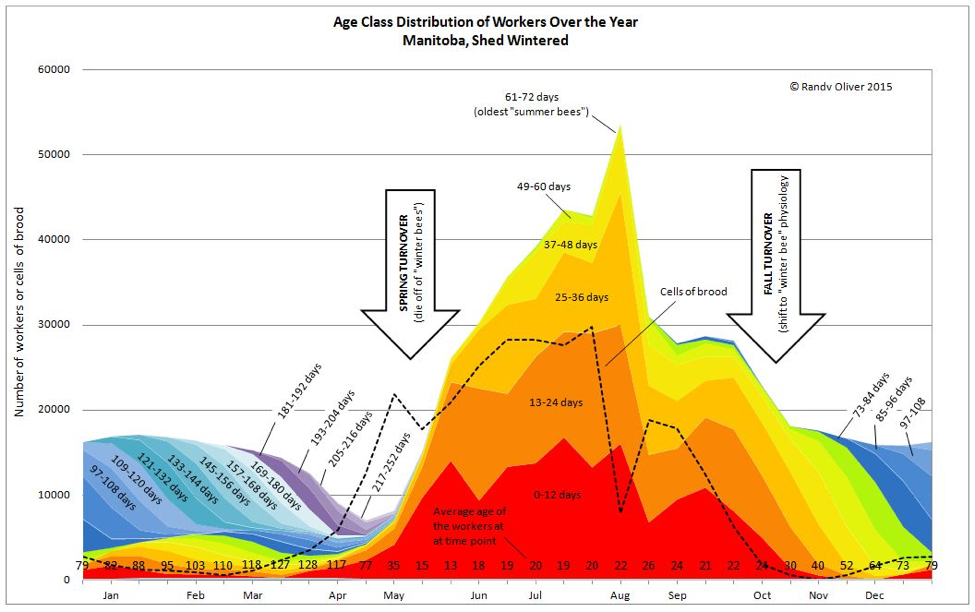
Think about it another way—becoming a queen is the default end result for any fully fed female larva. It is only by suppression of this “normal” development that a larva is turned into a worker [8], and only due to the inhibitory effects of pheromone(s) produced by the queen that that worker remains functionally sterile.
It seems then, that a female honey bee has the potential to live for a long time. A queen (reared from a worker egg) can live for years, despite her metabolically-exhausting task of clambering all over the broodnest while producing up to her body weight in eggs each day. And extended longevity doesn’t apply only to queens–workers commonly live for over 200 days during the winter [9]. Clearly, any female bee is born with the potential to live for a long time.
Practical question: so why do summer worker bees live for such a short time?
The Regulation Of Aging
There are two main explanations for the process of aging, or more properly, “senescence”:
- The slow cumulative damage caused by biological processes, environmental stressors, oxidative damage to tissue, and general wear and tear (we beekeepers in our Golden Years are unfortunately quite familiar with this concept).
- Innate “programmed senescence,” in which at some time point in an organism’s life, there occurs an innate epigenetic down regulation of tissue repair genes—thus preventing the body to continue repairing itself, thereby accelerating the onset of death.
For any organism, there is a tradeoff between the cost of body maintenance or rapid development and reproduction. Thus, species with an expected high risk of predation or starvation (think mice) tend to favor rapid growth at the expense of longevity. On the other hand, species with a high chance of survival (think Galápagos tortoises), invest in body maintenance, delay reproduction, but then eventually reproduce for decades. The tradeoff between maintenance and growth/reproduction is determined by which is most likely to result in the passing of that species’ genes into the next generation.
Class Insecta offers some extreme examples. Adult mayflies, which only need a day or two in which to mate and lay eggs, make no physiological effort to live longer than that. Compare that to the queen army ant, which can live for 20 years, during which she may lay 50 million eggs. Similarly, a female honey bee can develop into a short-lived summer worker that lives for little more than month, a long-lived “winter bee” (living over 200 days), or queen that can be productive for years.
The Honey Bee As A Model For Gerontological Research
I’m hardly the only one who has pondered the question of bee aging. In 2002, Gro Amdam and Stig Omholt published a seminal paper [10] that explained how and why honey bees regulate their longevity as an energy-saving mechanism, premised on the utilization of the yolk protein vitellogenin (Vg). They gently suggested that:
We think [this] paper substantiates the claim that an understanding of the regulatory anatomy of honeybee lifespan in evolutionary as well as proximate terms might be of considerable gerontological interest.
That turned out to be an understatement. Bee researchers are always scrambling for funding, but humans, ever in search of the Fountain of Youth, gladly throw money at gerontological research (the study of aging). Amdam followed with a series of papers, soon collaborating with Olav Rueppell and Rob Page Jr. [11]. The die was cast, and the honey bee had become a model organism for the study of the regulation of senescence (with the teaser that we could apply the findings to humans) [12]. Despite the fact that most of the studies were published in journals dealing with aging or theoretical biology, bee biologists benefited greatly from this research, and I thank Dr. Amdam for her continued studies.
Most of Amdam’s research has focused upon the yolk protein vitellogenin [13]. The honey bee superorganism, which consists largely of sterile female workers (thus having the ability to produce vitellogenin, but having no use for it), coopted this remarkable substance for other uses–as a currency of protein within the hive (via conversion into jelly), as a way of stockpiling protein (in the fat bodies), as an immune system component and antioxidant [14], as a regulatory hormone (an antagonist of juvenile hormone) [15], and for transgenerational immune priming from mother to daughter [16].
In short, the honey bee colony revolves around vitellogenin. So it shouldn’t come as a surprise that it also uses it as a regulator of senescence.
Practical application: an easily observable indirect indicator of the titer of vitellogenin in the colony is the amount of jelly being fed to early instar larvae [17].
Bee Female Phenotypes
We often hear of the two castes of female honey bees—queens and workers [18]. But Amdam [19] points out that there are actually four functional phenotypes (term: phenotype– the composite of an organism’s observable characteristics or traits, such as its morphology, development, biochemical or physiological properties, phenology, and behavior).
Those four phenotypes (or functional castes) can be defined by their tasks, as well as by their content of vitellogenin and closely correlated longevity:
- The reproductive queen caste; extremely high Vg, longevity up to 5 years.
- The diutinus (“winter”) bees; extremely high Vg, projected longevity up to 2 years [20].
- The nurses (and mid-age workers); high Vg, mean longevity of around 2-3 weeks.
- The foragers; negligible Vg, mean longevity 8-12 days.
The different longevities of the four phenotypes are closely tied to the degree of the colony’s nutritional investment in each, as well as the expected return on investment. Amdam explains:
[Natural] selection will in this case favor the survival of individuals that will make larger investments in others in the future; or equivalently, individuals that embody a larger investment of resources than others are under positive selection for survival.
Longevities of the queen, diutinus bees, and the foragers
The colony has tremendous expectations of the queen, as well as from the diutinus bees (to rejuvenate the colony in midwinter by drawing upon their stored Vg). So it’s to the colony’s adaptive advantage to keep the queen and diutinus bees safely ensconced and protected within the hive, enjoying minimal senescence.
Nurse bees, being responsible for pollen digestion and distribution, the production of jelly for the benefit of the queen and foragers, as well as the maternal care of the brood, also give a good return on investment, and thus enjoy somewhat delayed senescence.
Once a worker transitions to foraging, however, it is a different story. Although foragers are critical for bringing food to the hive, their remaining lifespan is limited at the very least by the irreparable wear and damage to their wings. From the colony’s perspective, any individual forager is a poor return on investment, so it doesn’t waste resources on it. Thus, prior to transitioning to foraging, the worker “slims down” by feeding any remaining vitellogenin in its body back to younger bees in the hive, thereby minimizing the removal of protein from the hive. As a result, that forager, even if it avoids environmental risks, wear and tear, will quickly succumb to lack of tissue maintenance.
Practical thought: although the strategy of saving risky behavior for its oldest members certainly works for the honey bee, I’m not sure that it’s the best idea for aging beekeepers.
As proposed by Rueppell [21]:
… it seems fair to conclude that aging in honeybees is not merely a collection of nonadaptive deleterious events that happen in the shadow of natural selection, but that natural selection has shaped the aging pattern to come under strict regulatory control that ensures improved allocation of resources at the colony level.
An important assumption: it’s likely safe to assume that the aging of each phenotype of bee is under “strict regulatory control.” Our question then, is exactly how each phenotype’s aging is regulated. Let’s start by figuring out how a worker “knows” whether it should become a nurse bee or a diutinus bee?
The Two Paths
Of interest is that the nurse-to mid-age-to forager transition is reversible if necessary (to a lesser degree once the transformation to forager is complete). But it appears (though not yet fully tested) that the transition to nursing is irreversible–once a nurse, a worker can no longer revert to being a long-lived diutinus bee. I doubt that this is due to chance.
If you take a look at Lloyd Harris’s data in Fig. 1 above, you can easily see (to the right) that diutinus bees (blue and violet shades) do not appear until broodrearing (dashed curve) ceases. And then they survive with little mortality until they initiate substantial broodrearing (to the left), at which point they immediately revert back to the “summer bee” bee survivorship curve (as though they had not already lived over a hundred days) [22].
Heather Mattila [23] also analyzed Lloyd’s data set, and concluded that:
Our survivorship curves suggest that worker mortality decreases (therefore worker longevity increases) as brood production declines in the fall; colonies in which brood rearing ceased earlier showed earlier shifts in worker survival.
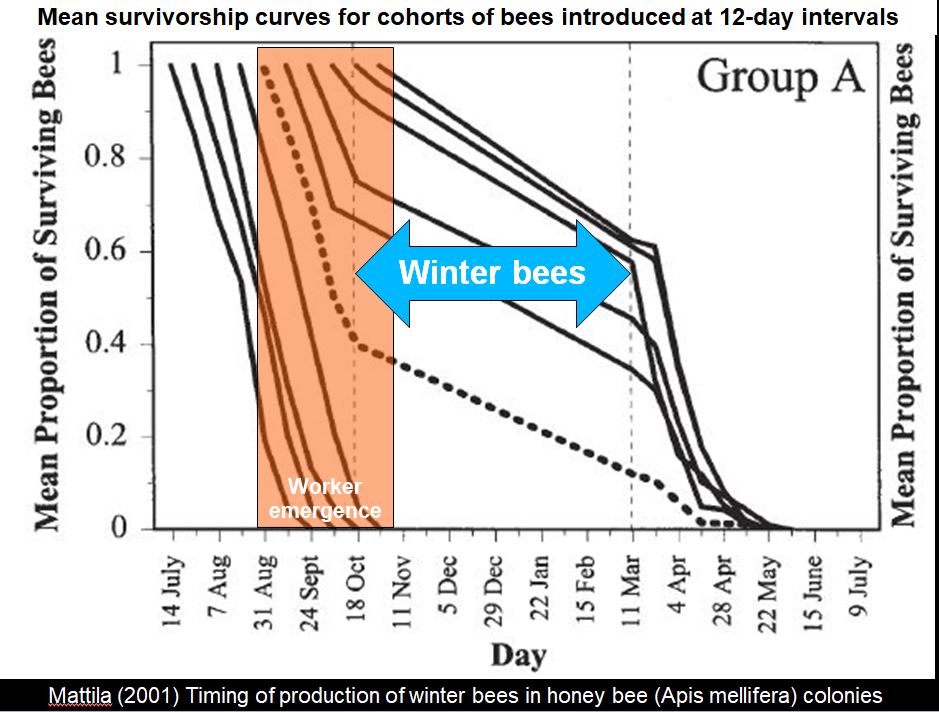
In the above graph from Mattila, I’ve overlaid with shading the time when winter bees are produced. The emergence date of each age cohort is where each line starts at the top of the graph. Only bees that emerged in the orange-shaded area survive the winter. Once they begin broodrearing in the spring, the start to quickly die.
She later confirmed this with a study of her own [24]. The remaining question (which Lloyd and I have long debated) is whether the abrupt shifts in worker longevity in spring and fall are due to an epigenetic shift having to do with the act of rearing brood, or are simply due to the stress of foraging. In other words, is it the lack of broodrearing that confers diutinus survivorship on those workers, or is it the lack of foraging?
To my mind, this question was largely answered by a well-designed study by the Amdam group [25], in which the rearing of brood alone caused workers to become short-lived bees. But of even greater interest was that the mere presence of artificial brood pheromone (without any actual brood being present) was enough to create this effect.
This was an arresting finding. I’ve written about brood pheromone (BP) acting as a primer pheromone [26]. It’s certainly plausible that upon emergence (Fig. 2), a worker’s fate is determined by the cue of whether brood pheromone is present or absent. This would constitute an elegantly simple regulatory system. A single cue would immediately inform the worker which of two developmental paths it should take. The cue of brood (and perhaps the scent of fresh pollen) would be clear signals that the colony was in growth mode, and the worker should start to develop its brood food glands (at the expense of longevity). A lack of brood pheromone would give the signal that the colony was in nutritional stress, and that the worker should take the path of going into conservation mode—storing all the protein it could in its fat bodies, and hunkering down for an indeterminate wait until conditions called for it to shift into broodrearing when pollen again became available. Beautiful!
 I created a chart to visually illustrate how this pheromonal regulation apparently works (Fig. 3). Keep in mind that this is only a proposed model (being vigorously debated even as I sent this article off to press).
I created a chart to visually illustrate how this pheromonal regulation apparently works (Fig. 3). Keep in mind that this is only a proposed model (being vigorously debated even as I sent this article off to press).
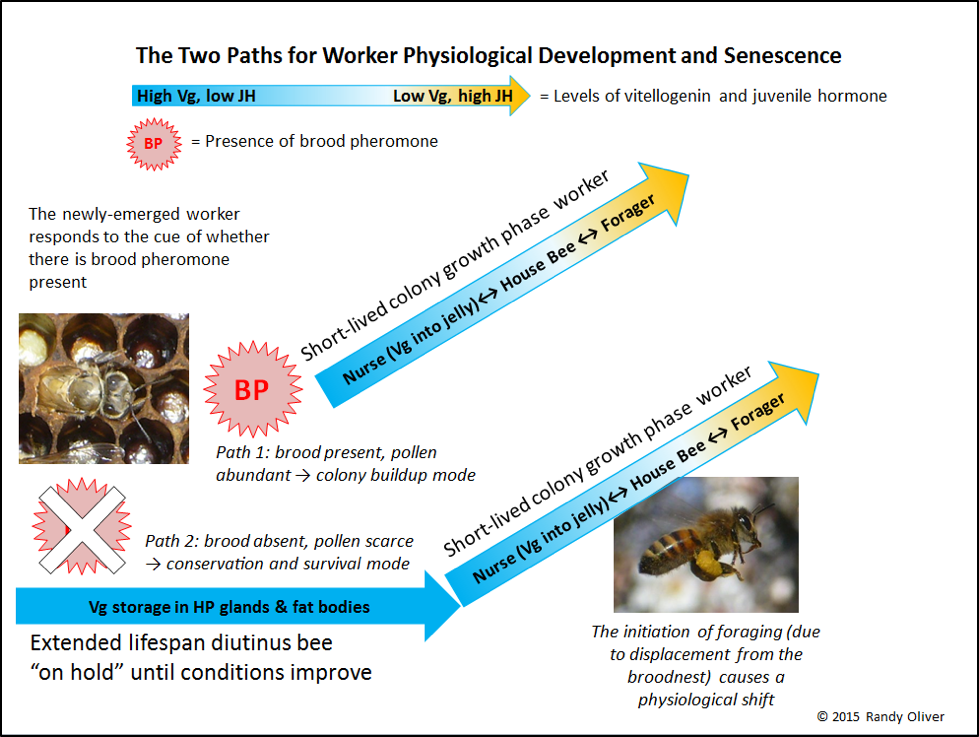
There is likely redundancy in this system (as there is in all bee regulatory systems). Besides the cue of brood pheromone, the odor of pollen, the amount of jelly fed to newly-emerged bees by the existing nurses [28], and other pheromones may well also play a role in the “choice” of path taken.
An alternative hypothesis: Lloyd Harris and I continue to debate the regulatory signals involved, and he disputes the above hypothesis. In his words [29], the absence of brood pheromone is probably not that important of a factor in switching bee longevity from short lived bees to long lived bees. I would put my money on a combination of lower ambient temperatures and queen mandibular pheromone effects on JH levels in bees. Perhaps both. The lower ambient temperatures bringing bees into closer contact with the queen where the effect of QMP can suppress JH levels and consequently foraging behaviour or simply a low temperature effect on JH production in workers. I prefer the combination effect as an explanation.
Update 31 Jan 2019: beekeeper Peter Somers from Salt Lake City succinctly summarized seems to be the best working hypothesis for the formation of “winter bees”:
Winter bees are produced gradually over a 60+ day period in late summer and fall. Because the queen is laying less and less with each passing week during this time of year, there would be a surplus of young bees… more than is required to rear the declining amount of brood. So it seems with each round of brood, a growing proportion of those new bees are relieved of both brood care and foraging, and gradually accumulate from late summer onwards to form the winter cluster. (Emphasis mine).
Updates 11/21/2020
Other findings:
de Groot
de Groot observed that workers produced late in the season, as the ratio of adult bees to brood decreased, emerged with higher dry weights and protein levels than workers reared during spring and summer. This could help to set them up to become diutinus bees.
de Groot also explained why “winter bees” would be expected to have higher body protein content:
However, there is no need to ascribe an eventual difference to additional pollen consumption in autumn. Just the suggestion that nurse bees in autumn, due to lack of larvae to feed, remain in the stage of maximum development during the entire broodless season would suffice to explain a possible difference in nitrogen content between nurse bees released from nursing duties and those exerting their nursing duties.
de Groot, A. (1953) Protein and amino acid requirements of the honey bee. Physiol. Comp. Oecol. 3, 197–285.
(Rueppell et al., 2007). Indeed, the foraging stage is ultimately linked to more risky tasks and thus to earlier
death (Neukirch, 1982).
Eyer
This recent study sheds new light upon “social regulation” of aging. The presence of young workers in the hive appears to signal older workers to transition to forager physiology, thus leading to their early death. On the other hand, if workers of nurse age do not sense the presence of replacement nurses, then they maintain their fat bodies, hunkering down as long-lived diutinus “survival bees.”
Our results show for the first time that the presence of young workers alone can significantly reduce the longevity of nestmates. .. In line with this hypothesis, the continuous emergence of young workers may accelerate the behavioral ontogeny of nestmates, i.e., workers may start foraging earlier in the presence of young adults, thereby decreasing their lifespan… Indeed, the foraging stage is ultimately linked to more risky tasks and thus to earlier death…
Eyer, M, et al (2017) Social regulation of ageing by young workers in the honey bee, Apis mellifera. Experimental Gerontology 87,(A): 84-91.
The science behind the shift to diutinus physiology is yet to be settled. It’s still unclear as to whether workers that have shifted to mid-age physiology can later transition to the diutinus state. Also unclear is the trigger(s) that cause the survivorship of the winter diutinus bees to drop off precipitously in early spring [30]. I’ve analyzed Lloyd’s data (as well as the weather record) and suspect (as I’ve shown in the chart above) that the switch from diutinus state to “normal” survivorship [31] occurred when emerging young bees displaced the elderly diutinus nurses from the broodnest, thus finally causing a drop in Vg and rise in JH [32]. However, to confirm whether temperature is involved, we need good data from colonies that have gone into diutinus state during summer dearth [33].
I look forward to seeing these competing hypotheses being experimentally tested.
Next
I’m out of room for now. Next I’ll continue with the regulation of aging in summer bees.
Acknowledgements
I thank Lloyd Harris, Gro Amdam, and especially Tom Seeley for their constructive comments on this manuscript. I also wish to express my appreciation of the other pioneering researchers (as well as their many students) who have guided my way in my attempts to understand bee behavior and colony homeostasis: Knut Schmid-Nielson, Bernt Heinrich, Paul Schmid-Hemple, Mark Winston, Karl Crailsheim, Yves Le Conte, Gene Robinson, Zach Huang, Rob Page, and Olaf Rueppell (apologies to those I overlooked).
As always, I could not research these articles without the help of my collaborator Peter Borst. And of course thanks again to Lloyd Harris for his data set and helpful discussion, without which I would never have written this series.
Notes And Citations
[1] Honey bees follow a generic model for social insects, as described by
Oster and Wilson (1978) Caste and Ecology in the Social Insects. Princeton University Press.
You can view their Figure 3-2, which is similar to the bee colony life cycle at http://www.antwiki.org/wiki/The_Ants_Chapter_3
[2] Under favorable conditions, bees in natural cavities may swarm more than once a season. A successful established hive, since it replaces its queen with a daughter each time it swarms, can live for many years, producing dozens of swarms in total (this used to be common prior to varroa).
[3] Such as for the maximization of the net energetic gain in a foraging trip per unit of energy spent:
Schmid-Hempel, P, et al (1985) Honeybees maximize efficiency by not filling their crop. Behavioral Ecology and Sociobiology May 1985, Volume 17, Issue 1, pp 61-66
[4] Szabo, TI & LP Lefkovitch (1989) Effect of brood production and population size on honey production of honeybee colonies in Alberta, Canada. Apidologie 20:157-163
[5 In The Wisdom of the Hive, p.248.
[6] Since there is little return on investment in drones whose genes have scant chance for survival in a colony founded during a pollen dearth.
[7] This is common in the arid West and Mexico. I had entire yards shut down broodrearing completely in August during this year’s drought.
[8] There are a number of mechanisms involved, including the orientation of the cell, the quantity and proportion of jelly and nectar fed, and epigenetic regulation by the substances royalactin and microRNAs in the jelly. And a recent paper suggests that the feeding of a small amount of pollen to worker larvae may also be a contributing factor.
Kamakura M (2011) Royalactin induces queen differentiation in honeybees. Nature 473: 478–483.
Guo X, et al (2013) Recipe for a Busy Bee: MicroRNAs in Honey Bee Caste Determination. PLoS ONE 8(12): e81661.
Mao, W, et al (2015) A dietary phytochemical alters caste-associated gene expression in honey bees. Science Advances 1(7) DOI: 10.1126/sciadv.1500795 Open access.
[9] The occasional worker is found to live a long time. Lloyd Harris recently wrote me that, “I marked/painted some more workers last September and to my surprise I had 1 worker bee that was still alive in mid-July. It lived at least 312 but less than 324 days. This is the longest lived worker bee that I have ever observed. It lived almost as long as those observed by Farrar.” Note that this worker was marked in fall, and was thus most likely a diutinus bee that never shifted to foraging the next summer.
[10] Amdam, GV and SW Omholt (2002) The regulatory anatomy of honeybee lifespan. J. Theor Biol 216: 209–228.
[11] Rueppell, Olav, GV Amdam, RE. Page Jr., and JR. Carey (2004) From genes to societies. Sci. Aging Knowl. Environ. 2004(5): 5. DOI: 10.1126/sageke.2004.5.pe5
[12] Lee, H-Y, et al (2015) Insects as a model system for aging studies. Entomological Research 45: 1–8.
[13] I’ve written about vitellogenin previously https://scientificbeekeeping.com/an-adaptable-workforce/
[14] Vitellogenin is the major carrier of zinc in bee haemolymph.
[15] Guidugli, K R, et al (2005). Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Letters 579: 4961-4965.
[16] Salmela H, Amdam GV, Freitak D (2015) Transfer of immunity from mother to offspring is mediated via egg-yolk protein vitellogenin. PLoS Pathog 11(7): e1005015. doi:10.1371/journal.ppat.1005015
[17] See my article When to Feed Pollen Sub https://scientificbeekeeping.com/when-to-feed-pollen-sub/
[18] Drones are sometimes referred to as being a “caste,” but in general, a more narrow definition of morphologically or functionally different subsets of females is usually used.
[19] Amdam, GV and RE. Page Jr (2005) Intergenerational transfers may have decoupled physiological and chronological age in a eusocial insect. Ageing Res Rev 4(3): 398–408.
[20] Omholt SW and GV. Amdam (2004) Epigenetic regulation of aging in honeybee workers. Sci. Aging Knowl. Environ Issue 26: 28. DOI: 10.1126/sageke.2004.26.pe28 Open access.
[21] Rueppell, et al (2004) op cit
[22] Refer to Fig. 3 in: Harris, JL (2010) The effect of requeening in late July on honey bee colony development on the Northern Great Plains of North America after removal from an indoor winter storage facility. Journal of Apicultural Research and Bee World 49(2):159-169.
I’d like to see a test of this by exposing diutinus bees to synthetic brood pheromone to see whether such exposure triggers the initiation of rapid senescence.
[23] Mattila, HR, JL Harris, and GW Otis (2001) Timing of production of winter bees in honey bee (I) colonies. Insectes Soc. 48: 88 – 93.
[24] Mattila, HR and GW Otis (2007) Dwindling pollen resources trigger the transition to broodless populations of long-lived honeybees each autumn. Ecological Entomology 32: 496–50.
[25] Smedal B, et al (2009) Brood pheromone suppresses physiology of extreme longevity in honeybees (Apis mellifera). Journal of Experimental Biology 212: 3795-3801. doi: 10.1242/jeb.035063
[26] See my series The Primer Pheromones, beginning with https://scientificbeekeeping.com/the-primer-pheromones-and-managing-the-labor-pool-part-1/
[27] Chart modified from: Omholt, Amdam (2004) op cit.
[28] Naiem E-S, N Hrassnig,, K Crailsheim (1999) Nurse bees support the physiological development of young bees (Apis mellifera L.). J Comp Physiol B 169:271–279.
[29] In his comments to a draft of this article, by permission.
[30] I’ve seen this sudden disappearance of the workforce occur in almonds just as the trees come into bloom and the bees begin intense foraging.
[31] Again, refer to Harris’ (2010) Figure 3, op. cit.
[32] There is no indication in Lloyd’s data that the rearing of brood by those diutinus bees during winter decreases their longevity. It was only when the temperature rose high enough for them to initiate foraging that their mortality rate suddenly increased. My explanation makes perfect sense, but that doesn’t mean that it’s true!
[33] As in the experimental “artificial winter” created in the oft-cited study by: Maurizio, A (1950) The influence of pollen feeding and brood rearing on the length of life and physiological condition of the honeybee. Bee World 31: 9–12.