
Understanding Colony Buildup and Decline: Part 12 – Late Summer Through Autumn

January 29, 2017
Understanding Colony Buildup and Decline: Part 12
Late Summer Through Autumn
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ are June 2016
I took a slight detour in my last article to address the “other bug” that we keep in our hives—the varroa mite. I plan to soon return to the mite. But first, let’s get back to the seasonal progression of the bee colony.
Although the colony typically declines in population during late summer, it takes advantage of late summer through autumn pollen flows in order to recruit a generation of workers that will carry it through the winter.
Late summer is a tough time for bees in the arid West where I keep bees. There is scant bloom, so colonies suffer from nutritional stress. And older queens begin to fail. Back in the day, colonies typically superseded failing queens, but since varroa, it seems (no supporting data) that more colonies go queenless and fail [1]. Oddly, there may also be a spike in swarming in late summer, sometimes due to varroa overload, but perhaps also from the process of late-season supersedure, or perhaps even as usurpation swarms [2].
And although I have made the case that it is to the colony’s benefit to dump excess workers when food is scarce, it is critical for good wintering for the colony to rear a generous round or two of brood in fall—producing well-fed workers that will become the long-lived “winter bees.”
Fall Broodrearing
For many years I moved my hives out of state to better pasture, not so much for the honey, but because they went into winter in much better shape for almond pollination the following February. But I eventually got tired of all the driving, and learned that I could get nearly as good results by feeding high-quality pollen sub to supplement the bit of local natural pollen. Feeding was more costly than migrating, but I preferred the lifestyle of staying closer to home. That said, there’s nothing like a good fall pollen flow (Fig. 1) to get your hives in good condition for winter!

Figure 1. For many years moved my hives to fatten up on the late-summer bloom of Rabbitbrush, here in bloom along an irrigation ditch in Nevada. Colonies wintering with a good reserve of beebread made from high-quality fall pollen require little or no stimulation to be in great shape for almonds.
I cannot emphasize enough the importance to the colony of good nutrition in late summer and fall. Those of you who live where it rains during the summer have no idea how lucky you are to have your Goldenrod and other fall bloom. I had to chuckle when I read the excellent research by Mattila and Otis in Ontario [3], in which they attempted to produce “pollen limited” colonies in autumn. The sheer amount of pollen they trapped from each hive per day, and the amount of beebread in the hives, would make a California beekeeper drool with envy!
Practical application: the last couple of rounds of brood reared in autumn are critical for colony winter survival, since it is those bees that will form the winter cluster. Successful beekeepers make sure that their hives are on either good natural forage, or spend money on pollen sub to ramp up broodrearing before it gets too chilly.
Practical application: my colonies eagerly consume pollen sub in fall. I’ve found that if a colony doesn’t consume its sub, that that’s an infallible indication that something’s wrong, and that that colony is unlikely to make it through the winter. Best to simply move it immediately to an isolated sick yard, and waste no further time or effort on trying to “save” it.
The Post-Brost Cessation of Broodrearing
Finally, in cold-winter areas, pollen becomes scarce or absent, and the colony ceases rearing brood. This typically occurs in late November or early December (but in subtropical climes, this is, contrarily, when the colony may reinitiate broodrearing after the dry summer pollen dearth, due to plants flowering when winter rains come).
Mattila [4] (analyzing Lloyd Harris’ data), showed how this cessation of broodrearing resulted in the production of “winter bees”—those bees that emerge after there are no more larvae to feed, shift their physiology into the long-lived diutinus state. And it is those bees that will hold down the fort for the winter.
The (healthy) colony (which for all it knows is living in a tree hollow) now has a problem. Bees which have served as nurses and foragers do not appear to be able to revert to the diutinus (long-lived) state. If those bees go into the winter cluster, and then the weather turns too cold for them to fly off to die, the cavity could soon start to fill with their corpses. It is to the colony’s advantage for those bees to fly off to die before it’s too late. And that’s just what they appear to do.
Dumping The Field Force
The data from both Lloyd Harris and from a large-scale Beeologics trial [5] in which I was involved suggest that no matter the strength of colonies in summer, (in the words of Harris) there is “a tendency for colony populations to converge towards a common numerical size.” This size appears aim toward a cluster of 9-10 frames in strength in December. Thus, stronger colonies appear to shed a greater proportion of excess bees.
Allow me to share a portion of my own data, for which I carefully graded a hundred hives for strength in October, and then again in December and January (99 survived). What was of great interest was the drop in strength relative to their October population (Fig. 2).
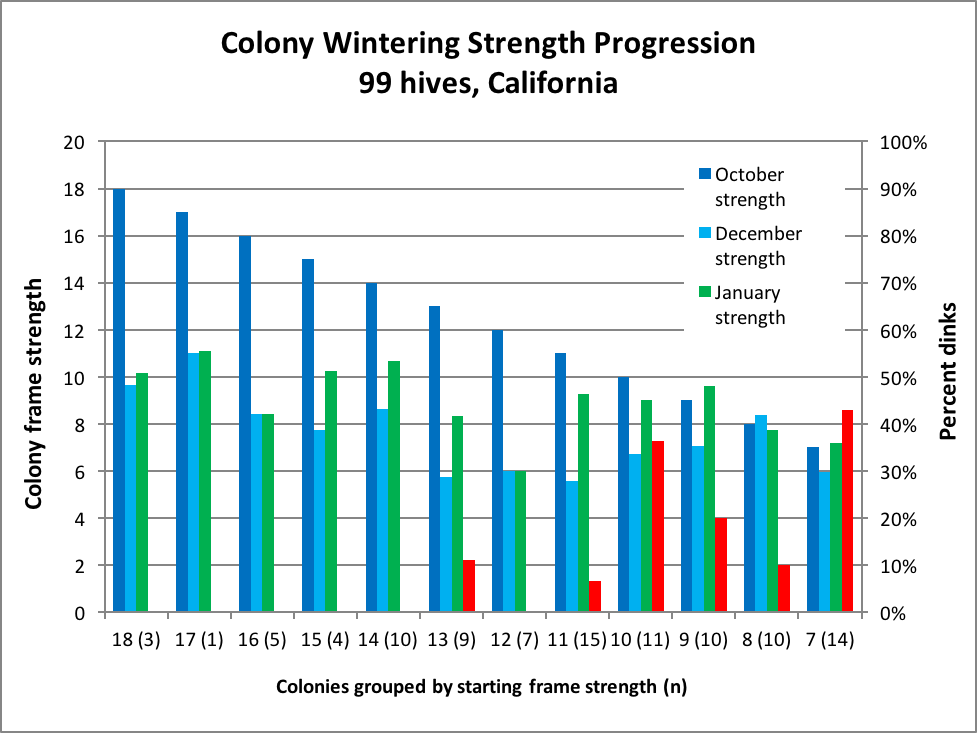 Figure 2. Look at the relative differences in height between the dark blue and turquoise columns. To help you understand this histogram, the “18(3)” means that there were 3 colonies that graded at 18 frames on October 27; they dropped, on average, to a bit over 9 frames in December, and rebounded only slightly by January 27. The “9(10)” means that there were 10 colonies that started at 9-frame strength; they dropped to 7-frame in December, and the red bar indicates that 20% of them (2 “dinks” out of 10) crashed before the January grading. Of those in the group that survived, they rebounded to an average of nearly 10-frame strength.
Figure 2. Look at the relative differences in height between the dark blue and turquoise columns. To help you understand this histogram, the “18(3)” means that there were 3 colonies that graded at 18 frames on October 27; they dropped, on average, to a bit over 9 frames in December, and rebounded only slightly by January 27. The “9(10)” means that there were 10 colonies that started at 9-frame strength; they dropped to 7-frame in December, and the red bar indicates that 20% of them (2 “dinks” out of 10) crashed before the January grading. Of those in the group that survived, they rebounded to an average of nearly 10-frame strength.
The above data was collected personally by me; it very closely matches the data set for 600 hives managed and graded by 12 different beekeepers (in which the red “dink” bars progressed in a more linear manner).
Three aspects of these data sets catch my attention:
- That stronger colonies tend to drop in strength to a greater extent in autumn than do healthy weaker colonies.
- That the weaker the colony in autumn, the greater the chance that it will fail by the end of January (there’s gotta be some reason that it was weak in the first place), and
- That colonies of any starting strength in autumn tended to grow toward 8-10 frames in strength by the end of January (the 99 colonies in my data set were treated identically; but I do not know how the other beekeepers treated theirs). Of interest is that an 8-10 frame colony is just large enough to fill the size of cavity preferred by swarms [6]—do our bees have an innate goal of wintering at that size?
And how about observation No. 1? Larger colonies lost roughly half their strength as they went into winter cluster. I’m hardly the only one who has made this observation. Strong colonies tend to dump a large portion of their population in the fall. As best I can tell is that the short survivorship of bees that have functioned as nurses and foragers that then leads to a rapid reduction in their population as they fly off to die.
Practical application: don’t be surprised by a sudden substantial drop in population of your strongest colonies in autumn—they’re only dumping excess “summer bees.” They’ll still maintain an adequate cluster of “winter bees” that can later initiate early broodrearing.
The “Fall Turnover”
The “fall turnover” is the replacement of the population of short-lived “summer bees” by long-lived “winter bees.” There’s no visible difference in the change, but those winter bees are physiologically distinct. Those “winter bees” are generated from the last generations of autumn bees that do not then engage in broodrearing or foraging.
Practical application: those emerging “winter bees” must have access to either stored beebread (or artificial pollen sub) in order to fully develop their fat bodies, which then extends their longevity.
The Effect of DWV and Nosema
The exit of those late-summer bees from the hive may well be exacerbated by infectious disease. Natsopoulou [7] found that infection by DWV (and to a lesser extent Nosema ceranae), accelerates the behavioral progression of workers, quickly moving them out of the broodnest and into foraging, thus reducing the worker’s overall longevity.
In my last article I covered why DWV (or one of the paralytic viruses) becomes such an issue in late summer and fall, due to varroa finally building to a level of prevalence in the brood so as to initiate a virus epidemic. Not only can that virus infection cause the rapid exit of “summer bees,” but if the all-important “winter bees” are infested by varroa during their development in that last round of broodrearing, there’s no way that they will be able to make it through the winter.
Practical application: the future “winter bees” need to be relatively free of varroa and DWV during their pupal development in order to be able to survive the winter. Many beekeepers wait until it’s too late before they hop on mite management in fall. Successful beekeepers work hard to get mite levels down prior to fall broodrearing.
But how about nosema? Below are data for monthly spore counts and prevalence of infected bees in 30+ colonies in one of my yards in the Sierra Foothills, taken monthly over the course of a year (Fig. 3).

Figure 3. I measured the prevalence (number of infected bees out of 10), as well as mean spore count of 25 bees, over time, in a group of 36 hives in my operation [[i]]. Note how nosema gains traction in fall as the temperature drops.
[i] The study actually ran from September 2012 through August 2013, but I reordered the dates to match the calendar year.
Surprisingly, though, at least in my relatively mild winter climate, there was no correlation at any time between nosema counts and either colony strength or survivorship. I suspect that in a colder climate, nosema would have a stronger effect. Nosema can’t be good for the colony, but it is still unclear as to whether best management practices for all areas call for treatment with fumagillin [9].
Update Jan 2017: A recent study from Uruguay [10] similarly was unable to confirm any correlation between colony strength and the degree of infection from N. ceranae, nor any benefit from treating with fumagillin. As with many other researchers, I’m having trouble figuring out just how much of a problem N. ceranae actually is. There was good research demonstrating the benefit of controlling N. apis over winter, but there appears to be little evidence (other than perhaps in Canada) of such benefit for ceranae.
Practical application: those colonies that are weak in autumn are likely weak for some reason. In my experience, in only a portion is that weakness due to a poor queen, since I often see those colonies quickly turn around and regain their strength in spring. This makes me suspect that weakness in fall often indicates that the colony is dealing with disease; and the weaker the colony, the more likely it is to fail over the winter.
We’ve now passed the “fall turnover,” and the colony can now take a brief break before it resumes broodrearing in anticipation of the upcoming spring.
Acknowledgements
I cannot begin to express my gratitude to my colleague Peter Borst, who not only helps me with researching the literature, but also in challenging me as I weigh competing explanatory hypotheses for what makes the bee colony tick.
notes and citations
[1] My bees get scant exposure to ag chemicals, and my combs are free of synthetic miticide residues, so I can’t blame the increased rate of queenlessness on either of those suspects. Or perhaps I’m just imagining it.
[2] See https://scientificbeekeeping.com/whats-happening-to-the-bees-part-2/
[3] Mattila, HR & GW Otis (2006) The effects of pollen availability during larval development on the behaviour and physiology of spring-reared honey bee workers. Apidologie, 37(5): 533-546.
Mattila, HR & GW Otis (2007) Manipulating pollen supply in honey bee colonies during fall does not affect the performance of winter bees. Can. Entomol. 139: 554-563.
[4] Mattila, HR, JL Harris, and GW Otis (2001) Timing of production of winter bees in honey bee (Apis mellifera) colonies. Insectes soc. 48: 88 – 93.
[5] https://scientificbeekeeping.com/winter-colony-losses/
[6] Seeley, T (1977) Measurement of nest cavity volume by the honey bee (Apis mellifera). Behavioral Ecology and Sociobiology 2(2): 201-227.
[7] Natsopoulou, ME, DP McMahon, D & RJ Paxton (2015) Parasites modulate within-colony activity and accelerate the temporal polyethism schedule of a social insect, the honey bee. Behav. Ecol. Sociobiol. DOI 10.1007/s00265-015-2019-5 Open access. The authors have an excellent general discussion on the honey bee adaptive response to parasite pressure.
[8] The study actually ran from September 2012 through August 2013, but I reordered the dates to match the calendar year.
[9] I suspect yes for Canada and no for areas in which there is flight during the winter. But it would also depend upon the stock of bees kept, and perhaps what the bees eat (e.g., whether the fall honey or pollen causes dysentery, etc.).
[10] Mendoza, Y, et al (2016) Nosema ceranae winter control: Study of the effectiveness of different fumagillin treatments and consequences on the strength of honey bee (Hymenoptera: Apidae) colonies. Journal of Economic Entomology 2016: 1–5.


