



Understanding Colony Buildup and Decline: Part 8 – The Main Honey Flow
Understanding Colony Buildup and Decline: Part 8
The Main Honey Flow
Randy Oliver
ScientificBeekeeping.com
First published in ABJ Oct and November 2015
The moment has now arrived for our honey bees to earn their name. Unlike other insects in temperate climates, the honey bee maintains an elevated body temperature throughout the frigid winter. It does so by forming a cluster and metabolically burning honey to produce heat. Thus, survival of the species depends upon the efficient gathering of nectar and its processing into honey for long-term storage.
The Crux Of The Season
At this point in time, our colony has successfully completed its spring turnover, perhaps passed its genes into the next generation via some lucky drones or by the successful establishment of a swarm, and then completed its linear buildup to maximum population. All that remains to do is to gather and store enough honey to make it through the late summer dearth and extended cold and flowerless winter [1].
This decisive moment for the continued survival of the colony is now dependent upon the bloom, the weather, and whether the colony can exhibit optimal efficiency in nectar collection and storage. The bees can’t do much about the first two, but they are nothing short of remarkable in their ability to perform the last.
An Amazing Superorganism
The superorganism that we call the honey bee colony can be compared to a warm-blooded mammal weighing about 12 lbs [2]—roughly the weight of a large cat or small dog. But the bee colony can pull off a trick that a dog cannot do—part of it can “dissolve away” from the main body and temporarily spread out over an area of up to 80 square miles to forage for food, coalescing back into the whole each evening. But such foraging comes at a cost–a bee burns precious energy while flying [3]. The rate of energy loss sets the limit of the foraging range to a maximum range of about 6 miles [4]. But a colony does not forage over that area equally; efficient foraging is all about focusing upon food source “profitability.” With that much potential pasture to exploit, each hive in an apiary could conceivably forage in entirely different areas. We’ll return later to how the colony makes its decisions.
Ideally, by this point of time the colony has reached maximum population (Fig. 1). Not only that, but that population consists predominately of young, healthy workers (note the average age of the population along the bottom line), typically not encumbered by parasites or disease, and in optimal condition to take full advantage of what nature has to offer. So let’s look at how the population structure breaks down, and see how the colony allocates that workforce to the tasks at hand.
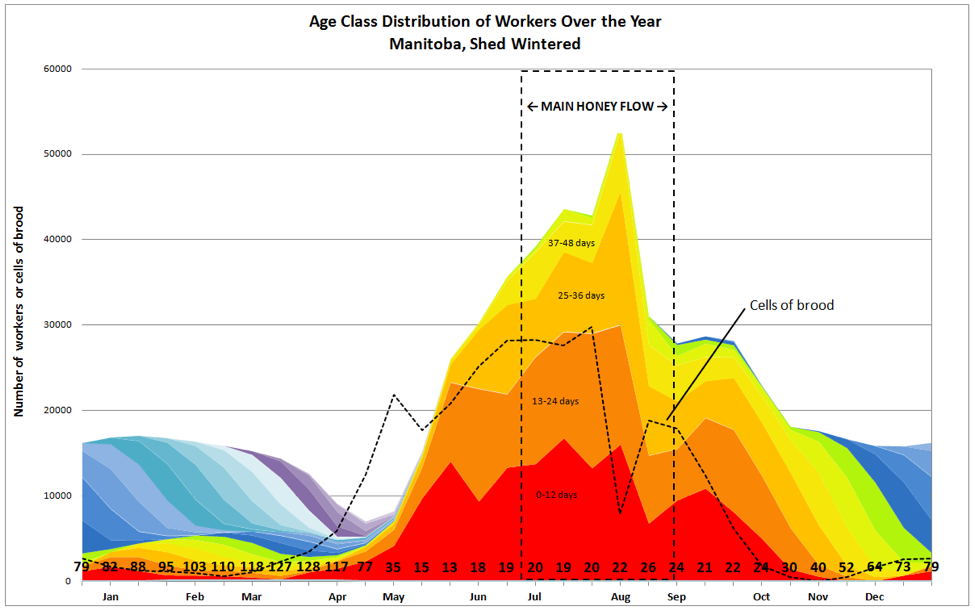
Figure 1. Note the large proportion of mid-aged bees and foragers (orange through lime green) at the start of the main nectar flow. And the curtailment of broodrearing (dotted line) as the call for “all hands on deck” prioritizes the storage of precious honey.
Practical application: keep in mind that the size of managed colonies greatly exceeds that of typical wild colonies, due to the much enlarged artificial cavity size [5]. We beekeepers thus use hardwired natural behaviors of the honey bee to our own benefit in order to get them to grow larger populations and store more honey than they would under natural conditions.
The Division Of Labor
Worker bees (similar to other social insects) during spring through fall follow a typical pattern of “temporal polyethism”—meaning that they change tasks (and their physiology) with age. They typically act as “cleaners” from days 0-3 after emergence, as “nurses” from days 4-13, as “mid-aged” general purpose workers from days 14-21, and then graduate to foraging for the remaining 11 or so days of their lives. Those figures are averages; workers are flexible, and if colony demand calls for it, they can shorten, skip, or extend any of those job commitments. It is this flexibility that allows the colony to take best advantage of ever changing opportunity at any time of the year.
Perhaps the most fascinating thing is how the colony “mind” works to quickly adjust the allocation of its workforce to changing conditions in order to maximize the profitability of its efforts. And it does so within the dark confines of the hive by individual bees communicating and coordinating their efforts via a combination of behaviors, feedback loops, pheromones, dancing, and vibrational signals. We’ll get to this, but first let’s return to…
The Broodnest
Up to this point in time, the colony has prioritized the broodnest. Farrar noted that “the honey flow is considered by the majority of beekeepers to be the most important external factor for stimulating a colony to a high level of broodrearing,” but points out that, at least with a strong flow, that this is a misconception:
There was no pronounced change in the rate of brood-rearing due to incoming nectar revealed by the data obtained during four seasons. There certainly was no stimulative effect and there were several periods where one might interpret an inhibiting effect due to the crowding of nectar into the brood nest.
The fact is that the honey bee colony faces an interesting dilemma when there is a strong nectar flow—should it focus upon maintaining the rate of recruitment (by ensuring that the broodnest remains “open” for egglaying), or should it go all out to collect and store the maximum amount of precious nectar (in order to avoid later starvation) while it’s got the chance? It’s pretty clear that nature has selected for the latter strategy.
Note in Figure 1 how the amount of brood tops out just as the main flow begins, and then suddenly plummets as the colony uses the broodnest for temporary nectar processing. One might ask, why would the bees do that? Wouldn’t that be nonadaptive?
Consider the selective forces that drove the evolution of bee behaviors. For most of the year, the main concern for the honey bee colony is to avoid starvation. In temperate climes, a colony may gain weight during only a few weeks out of the year (and loses weight the rest of the time) [6]. The only way the honey bee colony can expect to survive the winter is to take full advantage of those sporadic nectar flows, and to conserve the invaluable stored honey at all other times (remember that bees didn’t evolve with deeps of honey to draw upon). So when a serious nectar flow happens, the colony turns its focus to that, at the expense of broodrearing.
During a strong nectar flow, the mid-aged nectar receivers (and often pollen foragers) “take over” the broodnest and pack the cells with food. This of course prevents the queen from laying to her full capacity, and soon frees up young workers from nursing duties. This strategy allows the colony to focus upon nectar foraging, honey storage, and comb building.
It also has the benefit of reducing the number of hungry mouths to feed once the nectar flow ends. Consider this—it would be nonadaptive for a colony to forego the storage of precious winter stores in order to rear a generation of hungry workers that would emerge only after the flow was over. Better to quickly reduce the adult population immediately after the flow, which is exactly what happens.
Refer again to Fig. 1 to see how the curtailment of broodrearing in late July caused a subsequent rapid decline in the adult population after the main flow, as the workers that had worn themselves out by intense foraging quickly died off.
Practical application: the main determinant of the development of a busting colony for honey production is having a steady supply of pollen and nectar coming in well in advance of the flow. It is during that time that stimulative feeding and pollen sub can be of great benefit during periods of inclement weather or pollen dearth (as occurs in my area immediately after the end of fruit bloom). Immediately after a strong main flow, you can expect the colony population to greatly decline, which has serious implications in varroa management.
So let’s take a look at how the colony allocates its workforce during the main flow. This subject is wonderfully covered by Dr. Tom Seeley, who has dedicated his life to figuring out bee behavior. I highly recommend his book The Wisdom of the Hive [7], from which I drew much information for this article.
The Nurse Bees
The huge proportion of workers of nursing age becomes redundant as the amount of open brood decreases. This allows relatively young bees to fast track right to mid-aged (or even foraging) duties. So I’m going to skip right to those tasks.
The Mid-Aged Bees
Essentially, all workers over three days old who are not acting as either nurses or foragers can be considered as mid-aged (general purpose) bees. It is these bees that allow the colony to take full advantage of a strong nectar flow. And the larger the colony, the better it can do so. When I was investigating the limitations on spring buildup due to the necessity of the heat-generating “shell,” I created the chart below (Fig. 2), which suggests why larger colonies are able to store proportionally more honey than smaller hives.
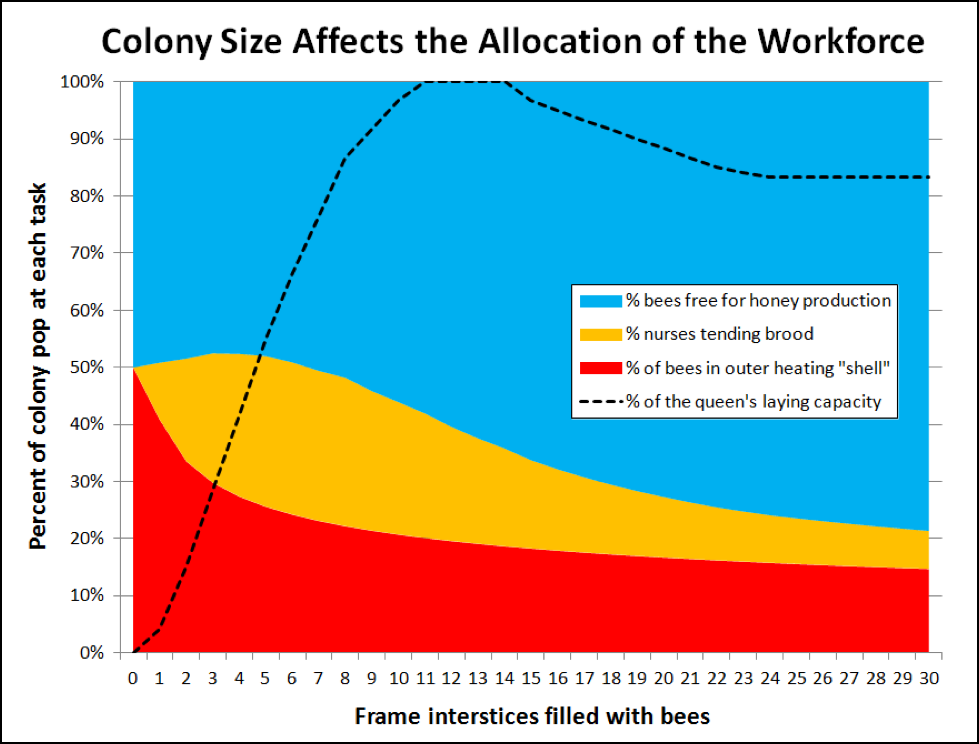
Figure 2. As a colony grows in size (from 0 to 30 frames of bees), the relative proportion of workers available for foraging and nectar processing (blue) increases substantially, which is why colonies of over 20-frame strength are the best honey producers. I created this theoretical chart for illustrative purposes only (my assumptions are at [[i]]); it likely overestimates the proportion of bees necessary for thermoregulation (red) during summer [[ii]].
[i] I assumed 2400 bees per interstice between the frames and a peak egglaying rate of 2000 eggs/day. I then calculated the volume occupied by bees (assuming an ellipsoid cluster), and calculated the percentage occupied in a ½” insulating shell. For the percent of nurses, I used the amount necessary to cover the open brood.
[ii] At this time the thermoregulation of the broodnest appears to shift from the insulating shell to far fewer “heater bees” in the broodnest itself, as described by Dr. Jürgen Tautz:
Tautz, J (2008) The Buzz about Bees: Biology of a Superorganism. Springer.
The linear decrease in the ratio of total population to sealed brood is well described in Farrar’s thesis.
Of course I needed to check whether the above chart reflects reality. Surprisingly, I was only able to find a single study that measured the percentage of a colony’s population involved in foraging. Danka [10] estimated that 6-10% (but up to 22%) of the population is out foraging in smallish colonies. I also asked Dr. William Meikle (ARS Tucson) to share some of his yet unpublished data, in which he tracked the variation in colony weight change throughout the day, which came up with similar figures. The higher figures match the “common wisdom” that up to a quarter of the colony goes out to forage.
So let’s do some math! A foraging trip on abundant local bloom takes about 30 minutes; and Seeley found that in nectar-choked hives a receiver bee could also take about 30 minutes for a processing cycle. This means that a colony may require as many receivers as foragers; so the blue area for smaller colonies should add up to about 50%, which supports the model. Now I’m curious to confirm these numbers in larger colonies next season [11].
Practical application: until colony strength approaches 10 frames, the queen is unable to lay to her full capacity (which limits the buildup rate of small splits). From that point on, the broodnest doesn’t get any larger, but the colony keeps building up a larger proportion of workers available for honey production. Thus one strong hive is able to out produce two weak hives.
Viewing a hive from outside, it’s easy to focus upon the foragers; but it’s the mid-aged nectar receivers inside that transfer that bounty to the combs. Thus, the colony must strike a dynamic balance (changing hour by hour, as well as day to day) between the number of returning foragers and the number of receiving house bees. In addition, those mid-aged bees also provide the jelly that the foragers need for body maintenance (since in slimming down for flight, foragers give up their ability to digest pollen or store protein).
We’ll return to how the colony balances the ratio of mid-aged bees to foragers in a moment, but let’s first give the foragers their due.
The Foragers
From the blue labor pool in the chart above, perhaps as much as a quarter of the workforce can act as foragers when called for. Think about it–the total body weight of the 50,000 bees in a strong colony works out to about 12 pounds. That means that there are perhaps only 3 lbs of bees bringing in up to 10 lb of nectar a day (over three times their body weight)—each forager returning with the equivalent of only one small drop of nectar per trip (Fig. 3)!

Figure 3. Beekeepers love to see returning foragers dragging their butts heavy with nectar. It takes more than 100,000 loads of nectar for the colony to make a 10-lb gain.
Consider that on average, a worker spends the first two thirds of her life in the dark and safe confines of the hive, only flying out occasionally to defecate. It’s to the colony’s benefit for an older worker to continue to hang out idly until there is a true demand for her as a forager. When that occurs, she can then promptly shift to the dangerous and exhausting work of scouring the countryside in the wind and bright sun, wearing herself out in a week or two. But these foragers (at least those that survive for a while) quickly learn by experience, become worldly, and exhibit relatively complex thinking. And they communicate among themselves and with the house bees.
Foragers don’t waste their efforts. If foraging is not profitable, they’ll save their energy by simply again hanging out motionless in the hive (Seeley refers to them as “unemployed foragers”), waiting for scouts or successful foragers to goad them into action.
Practical application (to the colony): Foragers remember when their favored flower patch produced nectar, and will save energy by waiting in the hive until that time. And although a forager exhibits “species fidelity” on any single trip, she may forage on other plant species at appropriate times of the day. In almonds the feverish foraging in the morning can drop to zero by mid afternoon, as the foragers conserve their energy rather than wasting it on dry flowers.
Roughly a tenth of foragers engage in scouting for new food sources (even fewer when resources are abundant; but up to a third when there is a lack of nectar coming into the hives. By extension then, larger colonies will likely have a greater number of scouts in the field than would smaller colonies. Along this line, Donaldson-Matasci [12] found that:
Honeybees in larger colonies were more effective foragers than those in smaller colonies, and this is mediated by their ability to communicate resource location via the waggle dance. Their advantage seems to be a result of being in colonies with greater information-gathering capacity: larger colonies may be able to locate rich resources more quickly, and perhaps also to select among them more effectively…This could indicate that the value of the dance communication system is highest when variation in resource quality is high, because that is when the ability to selectively exploit only the most profitable resources is most important
Not only that, but larger colonies foraged earlier in the day and their foragers returned with more nectar per trip.
Practical application: during a strong flow with copious nectar in the immediate vicinity of the apiary, all hives do well. But it’s a different story when resources are limited or when there is a wide variety of species with varying nectar quality in flower. Keep in mind that when resources are limited, there is intense competition between colonies. In well-stocked apiaries, weak colonies may be losing weight at the same time that strong colonies are putting on honey. Strong colonies have a leg up on those weaker—their scouts can better explore the territory and are more likely to locate profitable patches of bloom. The stronger colonies can then quickly exploit windfall discoveries due to their ability to quickly recruit their large reserve of unemployed foragers into action. It also appears that strong colonies will forage at greater distance than weaker colonies [13].
Scouts and other foragers excitedly share new discoveries. This is the huge advantage that honey bees have over solitary pollinators—similar to humans, information can be passed from experienced bees to younger bees. This is critical, since there is roughly a 10% overturn of the forager population every day. It’s of huge benefit to the colony that experienced foragers are able to pass learned information to the inexperienced.
The primary communication of information is via the famous waggle dance, from which recruits learn the direction and distance to the indicated patch of flowers. Now here’s where it gets especially interesting, as it involves the individual thinking ability of the bee. A forager bee apparently uses an internal calculus to determine the “profitability” of any food source based upon distance, effort required in collection, sugar concentration of the nectar, and overall abundance of nectar coming into the hive. She then dances accordingly–the more profitable the source, the more vigorously and repeatedly the returned bee dances, thus spurring recruitment of more foragers to that flower patch. A forager that is not especially excited about her food source may dance only weakly, if at all.
Practical application (to the colony): the waggle dance is used to inform other foragers as to the source of nectar. The vigor with which the dance is performed conveys the forager’s assessment of the profitability of the source.
But simply dancing about a new discovery is only the first step, since the waggle dance takes place only on the dance floor near the entrance, and there may not be enough of an audience. An excited forager will also range into the heart of the hive to rouse idle foragers or even young bees from the broodnest into foraging activity. For this she uses the “shaking signal.” Experienced foragers appear to use this form of communication each morning to rally the team.
Practical application (to the colony): the shaking signal is used to “wake up” idle foragers or to recruit younger bees into moving to the dance floor to see what’s happening outside. It is done mainly in the morning following the previous day’s good nectar flow, or whenever a forager discovers a new nectar source following a dearth or poor foraging weather. Overall, it conveys the message that the colony needs to put more foragers into action. This communication sets the initial pace of foraging for the day, and allows rapid recruitment of additional foragers should the need arise.
Regulation Of The Forager To Receiver Ratio
But there’s more. Since it takes roughly the same amount of time for a mid-aged receiver bee to process a load of incoming nectar as it did for the forager to gather it [14], that means that the colony must somehow strike a balance to maintain an even ratio between foragers and receivers. It would be moot to have an excess of nectar-laden foragers, since the colony as a whole would be inefficient at unloading them, and the opportunity of the bloom would be missed. Clearly, the colony needs some sort of feedback loop in order to maintain the proper equilibrium. It does so by several means.
Mid-age bees eventually progress to foraging at an average age of about 24 days, but may do so at as early as 3 days of age to as late as 65 days. The colony must somehow regulate the transition of younger bees to forager status–it would be of no benefit to the colony for there to be an excess of foragers, since they would quickly, and likely fruitlessly, wear themselves out.
The baseline balance is apparently struck by the inhibitory feedback of the pheromone ethyl oleate, produced by the foragers. They transfer this pheromone to mid-aged receiver bees via their trophallactic exchange of nectar; a constant titer then inhibits the receiver bees from transitioning to foraging. But should the ratio of foragers to receivers drop, the resulting reduction in pheromonal inhibition then allows mid-aged bees to transition to foraging [15]. The story then gets more interesting.
The colony places a very clever behavioral restriction on the foragers—although they directly deposit pollen into the combs themselves, they’re not allowed to do so with nectar. And this creates a balancing feedback loop. If a returning forager is mobbed at the door by eager receivers, this indicates to her that the colony is hungry for nectar, and she will then dance vigorously as above, and perhaps use the shaking signal to recruit more foragers. But if she has to search for a receiver bee to unload her, the less inclined she is to waggle dance, and she will dance with less enthusiasm, so as not to recruit additional foragers that would also be unable to unload their nectar.
But she doesn’t stop there. She’ll then go off the dance floor to recruit additional receiver bees—even nurse-age bees from the broodnest. She does so by applying the “tremble dance” to spur them into receiver duty [16]. In this way a forager can communicate the message to the colony that it had better recruit additional receivers into action to process what could well be an ephemeral source of nectar.
Practical application (to the colony): should a returning successful forager have trouble finding receiver bees to offload her, she will then use the tremble dance to recruit additional receivers.
Brilliantly, the colony squeezes a second use out of the tremble dance. If applied to a younger worker, it recruits her into foraging; but if applied to a dancing forager, it will cause her to freeze and stop dancing (acting as a “stop signal”).
Practical application (to the colony): what an efficient use of a single dance–if applied to a young bee, it means “switch to nectar processing;” if applied to a dancing forager, it gives the message “stop dancing, since we’re overloading the receivers. “
Hence it appears that the tremble dance functions as a mechanism for keeping a colony’s nectar processing rate matched with its nectar intake rate at times of greatly increased nectar influx. Evidently the tremble dance restores this match in part by stimulating a rise in the processing rate, and in part by inhibiting any further rise in the intake rate [17,18].
Wait, it gets even better! A forager can also use the stop signal/tremble dance to warn other bees of peril. Nieh [19] either pinched a leg of foragers (to simulate an attack by a spider) or exposed them to alarm pheromone while they were visiting a feeder. The threatened bees would then give the stop signal to prevent other foragers from dancing for that location or odor (this may also apply to pesticide exposure, below).
In summary, the honey bee colony, with no central command, utilizes an entire suite of overlapping informational, stimulatory, or inhibitory communication methods in order to most efficiently allocate the workforce to appropriate tasks. By doing so, tens of thousands of individual workers are able to continually adjust and coordinate their efforts in a seamless manner for the efficient exploitation of ever changing and ephemeral food resource opportunities.
Pesticides
In agricultural areas, the main nectar flows may come from monocultural crops or the weeds in the margins. Unfortunately, bees there are also often exposed to insecticides. In areas where there are a great diversity of crops (such as in the California Central Valley), this forces beekeepers to play “pesticide roulette” (Fig. 4).

Figure 4. When I drove into this field to take a photo of bees working sunflowers, the smell of recently-applied insecticide to the tomatoes in the foreground was overwhelming. This patchwork of crops, each with its own pesticides, can create a tough environment for bees.
Due to the rapid turnover of the forager population, a strong, healthy colony can generally shrug off a one-time hit to its field force from a fast-acting, short residual insecticide spray, provided that those bees die outside the hive and do not contaminate the nest’s pollen and honey stores. Weaker colonies, or those stressed by poor nutrition can’t handle it as well. And two successive hits can be devastating.
Of even greater concern are the legacy effects from pesticide residues carried back by foragers and stored in the beebread and combs. This is an extremely hot topic these days, often muddled by lack of good data, rigid dogma, and outright misinformation [20]. I’ve discussed this at length in other articles.
Practical application: although the colony may be able to replace its field force following a spray kill, it may lose the “learned knowledge” of that field force. Experienced foragers apparently contribute greater nectar income to a colony than do new foragers (Fig. 5).

Figure 5. In a study by Dukas and Visscher during a period of scarce resources, foragers took a few days to learn the ropes–hitting peak efficiency about midway through their foraging lifespan (and doubling their take per day). It appeared that bees learn rapidly during their first days of foraging, and then physically wear out toward the end of their lives. Interestingly, the older bees tended to bring back larger nectar loads, but spent more time per trip in doing so (Data reworked after Dukas [[i]]).
[i] Dukas, R & PK Visscher (1994) Lifetime learning by foraging honey bees. Animal Behaviour 48(5): 1007–1012.
Storage Of Honey
The multipurpose mid-age bees not only act as nectar receivers and processors, they also fan, repair, guard, and produce wax and build comb. The initiation of wax production is triggered in some yet undefined manner by an excess of nectar entering the colony relative to the amount of comb available in which to store it. The beekeeper can easily observe when the mid-age bees’ wax glands are in production by the presence of “white wax” being added to existing darkened comb (Fig. 6).

Figure 6. My sons and I need to draw out about 5000 deep sheets of foundation a season in order replace sold combs and make increase (and do this during California’s drought). Thus we keep a close eye out for white wax, which is our cue that the colony is ready to draw comb.
One of the most common questions by beginning beekeepers is “how can I make my bees draw foundation?” The answer is, “you can’t make them do anything–they will decide for themselves when the colony requires wax production.” And that occurs when the mid-aged bees have trouble finding empty comb in which to store nectar (or syrup). Only then will their wax glands kick in; at which point they’ll start to entertain the notion of drawing foundation.
I long ago learned that if I placed a deep of foundation upon a strong single at the start of a serious nectar flow, that the bees would draw comb and fill it with honey so quickly that the queen never got a chance to lay a single egg; they’d fill the box wall to wall with honey in 10 days. Knowing this, I ran an experiment a couple of years ago (not yet published) on drawing foundation in a similar manner by feeding syrup.
At the end of the main flow, I shook the bees from the top boxes of ten strong doubles down into their lower (now single) brood chambers, and then added a deep of foundation. I then fed syrup via two 1-gallon chick waterers set on the top bars (so as to not limit the rate of consumption). To my incredible surprise, in every hive, the bees stored the first 6 gallons of 1:1 syrup in the bottom box before they would even think about drawing any comb–it seemed like the first 6 gallons simply disappeared down the drain! But by that point they’d filled nearly all the empty cells in the lower brood chamber, and very quickly started working in the new box, readily drawing foundation on the next 6 gallons (yes, 12 gallons total, yet still the upper boxes were not nearly filled with processed syrup).
Practical application: it takes a heckuva lot of nectar or syrup to draw foundation, and the bees won’t begin until they’ve already filled every existing drawn comb. But the moment that you start seeing white wax, they can draw comb amazingly fast. The thing that we hate the most is when we underestimate a flow and let colonies plug out with white wax—that means that we’ve worked all year, and then missed the boat at the critical moment by not supering up in time.
A mistake that beginners frequently make is to think that your bees will recognize your own mental image that that super of foundation should be converted into comb. Unfortunately, your bees do not share that vision. To them, a super of foundation is simply empty space in the wrong place; unless the mid-aged bees are already producing white wax, they are not going to start working up there until they’ve filled every cell below the honey band at the top of the hive.
Practical application: keep in mind that the innate behavior of the mid-aged receiver bees is to store nectar immediately around and above the brood in already-formed comb, not in some far-off artificial supers placed high in the cavity. The bees will not cross the honey band and queen excluder to draw that foundation until they’ve filled every single empty cell of drawn comb below the super with honey. It’s easiest to draw foundation directly above the broodnest during a strong nectar flow, or even higher up in the hive once the bees have already initiated the production of white wax (so long as the flow continues).
Coming
The next installment of this series will cover the post flow decline in population and resulting impact of the varroa/virus complex.
Aknowledgements
I’d like to dedicate this article to Dr. Tom Seeley, whose lifetime of research has elucidated so many aspects of bee behavior. Not only that, but Dr. Seeley has been inspiration and mentor to a whole generation of apicultural researchers that have enjoyed the benefit of his tutelage.
Of course I’d like to express my appreciation to Lloyd Harris for his data set and assistance, and my friend Peter Borst for his help in scouring the literature.
Notes And Citations
[1] Those of you lucky enough to enjoy bloom during the winter can just keep it to yourselves.
[2] 50,000 bees at 110 mg ea works out to 12 lbs.
[3] Although Eckert calculated that it was energetically inefficient for bees to forage at greater than 4 miles, Beekman measured 10% of actual foraging flights in the heather moors at greater than 6 miles. Dr. Jerry Bromenshenk showed me a striking video of flying bees filmed in infrared. Each bee is a glowing little furnace converting sugar into heat energy.
Eckert, JE (1933) The flight range of the honeybee, J. of Agricultural Research 47(8): 257-285.
[4] Beekman, M & FLW Ratnieks (2000) Long-range foraging by the honey-bee, Apis mellifera L. Functional Ecology 14: 490–496.
[5] In nature, and by preference, honey bees are evolutionarily adapted for cavities of about the same volume as a single Langstroth deep.
Seeley, TD (1977) Measurement of nest cavity volume by the honey bee (Apis mellifera). Behavioral Ecology and Sociobiology 30(2): 201-227.
Seeley, TD & RA Morse (1978) Nest site selection by the honey bee, Apis mellifera. Insectes Sociaux 25(4): 323-337.
[6] Seeley, T.D. (1995) The Wisdom of the Hive. Harvard Univ. Press; refer to his graph at https://scientificbeekeeping.com/the-economy-of-the-hive-part-2/
[7] Seeley (1995) ibid. Another good review, from which I also took much information, is in The Biology of the Honey Bee by Dr. Mark Winston.
[8] I assumed 2400 bees per interstice between the frames and a peak egglaying rate of 2000 eggs/day. I then calculated the volume occupied by bees (assuming an ellipsoid cluster), and calculated the percentage occupied in a ½” insulating shell. For the percent of nurses, I used the amount necessary to cover the open brood.
[9] At this time the thermoregulation of the broodnest appears to shift from the insulating shell to far fewer “heater bees” in the broodnest itself, as described by Dr. Jürgen Tautz:
Tautz, J (2008) The Buzz about Bees: Biology of a Superorganism. Springer.
The linear decrease in the ratio of total population to sealed brood is well described in Farrar’s thesis.
[10] Danka, RG, et al (1986) Foraging population sizes of Africanized and European honey bee (Apis mellifera L.) colonies. Apidologie 17(3): 193-202
[11] It’s a relatively straightforward process to measure—weigh the hive during full flight, and then shake all the bees from the combs and weigh again to determine the weight of the bees in the hive. Repeat the process at the end of the day when all the bees have returned to determine the full colony population. The difference would be the amount of foragers.
[12] Donaldson-Matasci, MC, et al (2013) Bigger is better: honeybee colonies as distributed information-gathering systems. Animal Behaviour 85: 585-592.
[13] Beekman (2000) op. cit.
[14] Seeley observed processing times ranging from 6 to 28 minutes. Winston states that nectar foraging trips run from 30-80 minutes. Thompson measured 50% of the flights on canola taking 10 minutes.
Thompson, H, et al (2015) Thiamethoxam: assessing flight activity of honeybees foraging on treated oilseed rape using rfid technology. Environ Toxicol Chem., Accepted Article • DOI: 10.1002/etc.3183
[15] https://scientificbeekeeping.com/the-primer-pheromones-and-managing-the-labor-pool-part-3/
[16] See Seeley (1985) op cit.
[17] Seeley TD (1992) The tremble dance of the honey bee: message and meanings. Behav Ecol Sociobiol 31:375-384.
[18] Nieh, JC (1993) The stop signal of honey bees: reconsidering its message. Behavioral Ecology and Sociobiology 33(1): 51-56.
There are likely additional modes of communication to be discovered, such as the “piping signal” suggested by Thom, C, et al (2003). Worker piping in honey bees (Apis mellifera): The behavior of piping nectar foragers. Behav. Ecol.Sociobiol. 53: 199–205.
[19] Nieh, JC (2010) A negative feedback signal that is triggered by peril curbs honey bee recruitment. Current Biology 20(4): 310–315.
[20] https://scientificbeekeeping.com/a-review-of-dr-lus-paper-on-neonics-in-massachusetts/
[21] Dukas, R & PK Visscher (1994) Lifetime learning by foraging honey bees. Animal Behaviour 48(5): 1007–1012.