



What’s Happening To The Bees? – Part 6: Mitotypes, Genotypes, And Tradeoffs In Fitness
CONTENTS
And What’s This Got To Do With Bees?
Nature Doesn’t Care About Pedigree
What’s Happening to the Bees?
Part 6: Mitotypes, Genotypes, and Tradeoffs in Fitness
Originally Published in ABJ Sept 2014
Randy Oliver
ScientificBeekeeping.com
In my last article, I showed how greatly the genetics of wild-living honey bee populations differed from those of managed populations. So let’s take a look at the processes that have led to that differentiation, and the consequences. But first, I should review some concepts involved in the selective breeding of bees.
In business or sports, success is all about being “better” than the competition. And so it is in nature, with each honey bee colony vigorously competing against the competition (other colonies) for resources, including nest cavities. And by supplying ideal nest cavities, early beekeepers gave “kept bees” a leg up on the competition. But there is no such thing as a free lunch—in nature, any advantage typically comes at a cost. Evolution is generally a zero sum game; by becoming better at any one thing (emphasizing one trait), a species must give something up in return (losing the ability to do something else as well).
What Is “Fitness”?
Nature mindlessly selects for the most fit of each species by the process of continually weeding out the less fit. But just what do we mean by “fit”? Here’s the key thing to keep in mind:
Nature does not favor specific traits nor reward success. Nature only penalizes failure. Those left after penalization are called survivors, or “more fit.”
Any species, race, or breed of any plant or animal is alive today only because it was more fit than the competition in the never ending (and ever changing) process of natural selection.
However, Nature’s definition of “fitness” may not necessarily be the same as that of man’s. When a species enters into a symbiotic relationship with mankind, man can change the definition of “fitness.” “Fitness” is a relative concept which is completely dependent upon the selective pressures exerted upon the species by its local environment. Man has the ability to drastically change those selective pressures.
For a number of wild animal species that live in proximity of human habitation, the results of man’s recent selective pressure has been that the definition of “fitness” now includes a strong fear and avoidance of being within gunshot range. On the other hand, several of species of plants and animals have profited from a symbiotic relationship with humans, as humans facilitate the realized niche of those with desired traits. For some of these, this symbiosis has been wildly successful.
For example, a single species of plant, Zea mays (a formerly obscure species of Mexican grass), with considerable human facilitation, now outcompetes all other vegetation on some 625,000 square miles of the Earth’s surface (an area nearly equal to the entire land surface of the 12 states of the American Midwest). Maize could certainly now be considered to be a highly successful species. Yet if humans were to abruptly withhold their support, maize would go extinct within a few years, unable to survive without human facilitation of its niche (the ancestral grass, teosinte, would continue to survive [1]).
How could the clear success of such a species have come to the point that it hangs from the thread of human husbandry? C’mon, how long do you think Pomeranian dogs, Holstein cattle, or Cornish Cross broiler chickens would survive as breeds in the wild? When we breed for certain traits favored by humans (selective breeding), the zero sum game extracts a cost–a species loses in turn fitness traits for hardiness and adaptability to the challenges of life in the wild.
In the case of maize, we’ve bred out a critical allelochemical [2] that the ancestral stocks produced to deter insect pests, made the plants more dependent upon water and fertilization, and bred for seed heads that are now digestible by ruminants and not conducive to self propagation. And of even greater concern, we’ve bred out natural genetic variability in the domesticated stocks, which greatly hampers their ability to adapt to changing environmental pressures [3].
And What’s This Got To Do With Bees?
In my previous article in this series, the pie charts of survivor matrilines of honey bees in feral colonies clearly showed that those lines of bees that we select for in managed populations are not necessarily the lines that demonstrate fitness in the wild. With the added parasite pressures of the varroa/virus complex and Nosema ceranae in the last couple of decades, few managed stocks survive for more than a couple of years without human intervention. Yet genetic analysis shows that some unbroken bloodlines of feral bees have been able to persist in the wild for a great many years.
Practical application: I, for one, would like to understand how those feral stocks have managed to survive, and whether we can apply that knowledge, and use those genetics, to breed healthier and hardier managed bees (I’m sick of expensive mite treatments and supplemental feeding). But before you start shelling out cash for specific bloodlines of bees, there is something you should understand about genetic testing.
Mitotype vs Genotype
Let me state that I am no geneticist, and the arcane terminology of the field gives me a headache. But there’s something that I’d like to make clear about the pie charts of mitochondrial DNA (mitotype) inheritance that I included in the last article. Mitotypes are useful for tracking matrilines, since they can only be passed from mother to daughter. But it is nuclear DNA (genotype) that codes for the form, color, behavior, and disease resistance of bees. The two types of DNA are largely independent [4], meaning that although mitotype is useful for tracking pedigree, it is mostly genotype that is involved in selective breeding for observable characteristics.
Let me give you an example. Dogs were bred from wolves, and differ in less than 1% of their genes. Each dog breed started from a very small founder population, which led to each breed being considerably inbred. Despite that fact, several dog breeds contain multiple mitotypes. My point is that we breed for observable (phenotypic) traits (coded for by genotype), not necessarily by matriline (determined by mitotype)—when you cross two matrilines, one mitotype is lost forever, whereas roughly half of each genotype is carried on in the cross. In both nature and in breeding, there may be some degree of introgression of both nuclear and mitochondrial genes back and forth from one population into another (Fig 1).
It gets even more complicated. Some genotypes appear to work only with some mitotypes (as an analogy, Chevy parts may not work well in a Ford). For example, the M lines (Western European dark bees) are fairly closely related to the A lines (African). Anyone who remembers the hot “German black” bees that used to be common as ferals can appreciate the similarities in their behaviors (although British beekeepers breed gentle M line stocks). The A and M lines can more readily swap nuclear alleles than either can with the C (Italian/Carniolan type) lines [5].
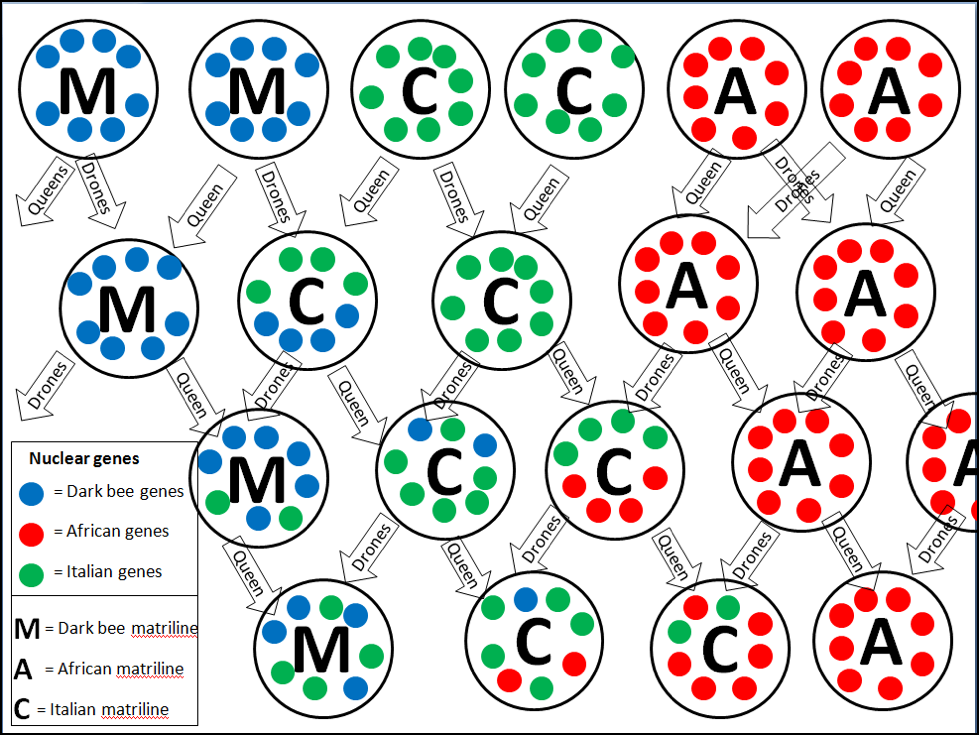 Figure 1. Beekeepers seem to prefer pictures with bright colors as opposed to textual explanations. So I created a hypothetical diagram of the introgression of nuclear genes [[i]] into different matrilines of open-mated purebred queens of three different races over three generations. Each large circle represents a “composite queen” of all mating possibilities [[ii]], with her mitotype indicated by letter, and nuclear genotype by colored dots. Note how mitotypes can only be passed mother to daughter (upper 4 dots), whereas nuclear genes can also be transmitted by drones (lower dots). In the end, a queen’s mitotype and genotype may be considerably different.
Figure 1. Beekeepers seem to prefer pictures with bright colors as opposed to textual explanations. So I created a hypothetical diagram of the introgression of nuclear genes [[i]] into different matrilines of open-mated purebred queens of three different races over three generations. Each large circle represents a “composite queen” of all mating possibilities [[ii]], with her mitotype indicated by letter, and nuclear genotype by colored dots. Note how mitotypes can only be passed mother to daughter (upper 4 dots), whereas nuclear genes can also be transmitted by drones (lower dots). In the end, a queen’s mitotype and genotype may be considerably different.
[i] Actually, various alleles of those genes. In this diagram, I arbitrarily used 8 dots to represent the degree of potential allelic diversity.
[ii] In reality, a queen only passes half of her nuclear alleles to an egg, so individual queens would not pass all the alleles shown.
In the above graphic, the purebred founders had nuclear genes attuned to their particular mitotype (all Ford parts in a Ford), and would likely have exhibited fitness. Hybrid [8] crosses are often less fit, since some mitotype/genotype combinations don’t work well (Chevy parts in a Ford), so not all of these hypothetical hybrids would have actually survived. But occasionally hybrids do gain in fitness—the Africanized bees of the Americas, although predominately of A line scutellata mitotypes, have incorporated a substantial proportion of European nuclear genetics [9].
Practical application: you can’t tell the genetic mixture of matriline and genotype by simply looking at a bee [10,11], or by her maternal heritage. In the graphic above, for example, the workers of the C mitotype queen to the lower right would almost certainly be highly defensive, due to the predominance of the African-type nuclear genes that code for behavioral traits.
Nature Doesn’t Care About Pedigree
Nature only selects against the less fit, and doesn’t give a darn about one’s pedigree. In natural selection, whatever works, works. Yet, many breeders focus upon maintaining unbroken matrilines (a number of European bee breeders maintain meticulous records of the pedigrees of their bees [12]). But as pointed out by Keoniger [13],
[M]ales contribute to the fitness of their offspring as much as females do.
Practical question: what are the respective relative impacts of the genetics carried by either drones or queens in nature or in breeding programs? Specifically, to what extent does a breeding program benefit from the contribution of the drone mother colonies or from matings with feral drones?
This may be a technical subject to the casual beekeeper, but has important practical application as we try to breed for bee stocks with better survival characteristics, especially with so many now attempting to propagate “survivor stocks.” I’ve read quite a number of studies on bee genetics, and am trying to sort out the relative contributions of mitotype and genotype to breeding programs [14].
In bees, there are no true “fathers” or “patrilines,” since the genetics of any drone come solely from its mother, and then every sperm produced by that drone is a genetic clone of the drone itself. And since sperm rarely contribute mitochondria to a fertilized egg, genetic mitotyping tracks only unbroken mother lines.
On the other hand, each female bee receives equal amounts of nuclear genetics (allelic forms of any gene) from both its parental drone and queen. And although the mitochondrial genetics of a colony are determined solely by the queen, the phenotype and fitness of that colony is affected by the nuclear genetics of the 10–40 patrilines of workers in the colony (each carrying in common half their nuclear genes from the queen, and half from their respective sires) (Confused? Perhaps Fig. 2 will help).
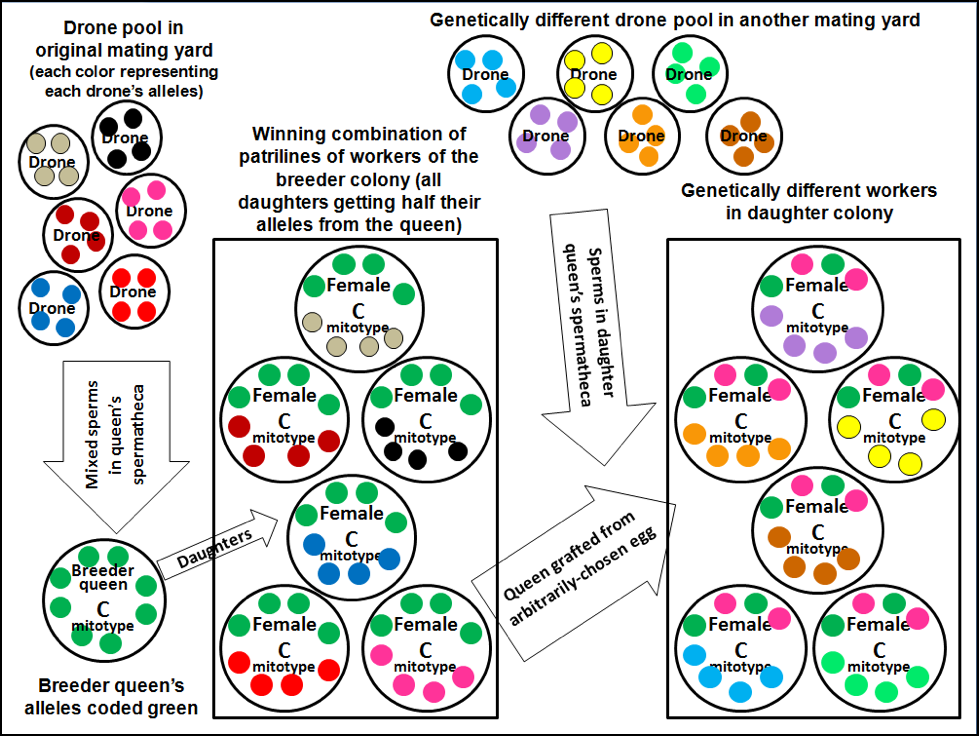 Figure 2. In this extreme (and even more colorful) scenario, every drone carries a different allele for each of the four genes illustrated (color coded by drone) [[i]]. Note how an open-mated breeder queen from a purebred (all green) line may produce a winning combination of genetically diverse workers. But when you graft a queen from that perfect colony and mate her out in a different yard, the resulting colony, although of the same queen line, may be substantially different genetically.
Figure 2. In this extreme (and even more colorful) scenario, every drone carries a different allele for each of the four genes illustrated (color coded by drone) [[i]]. Note how an open-mated breeder queen from a purebred (all green) line may produce a winning combination of genetically diverse workers. But when you graft a queen from that perfect colony and mate her out in a different yard, the resulting colony, although of the same queen line, may be substantially different genetically.
[i] Remember that the haploid drones carry only one copy of each gene, whereas the diploid queens and workers carry two copies, and potentially two different alleles.
Practical application: it’s difficult to propagate open-mated purebred lines of bees in a mongrel population such as we have in the U.S. (not that that is necessarily a bad thing). In the example above, the beekeeper grafted a daughter from a “dream colony,” the stellar performance of which was due to the perfect team of sister groups of workers (patrilines sired by different drones). But that daughter queen only carried half the alleles of the parent colony as a whole, and the daughter colony only a quarter of the (green) alleles of the original queen mother. This is a common frustration in bee breeding.
It’s abundantly clear that the nuclear genetics of drones may contribute substantially to the performance of their offspring, as evidenced by the often strong (positive or negative) effect of hybridization between a queen of one stock and a drone from another. Yet the distribution of matrilines in my aforementioned pie charts strongly suggests that some matrilines survive better in the wild than do others. I do not know why matriline appears to be so important.
Practical question: What can we learn from practical experience and studies to date? The data are conflicting.
Of interest is the famously successful invasion of the hybrid Africanized bees into the Americas, which have largely or completely displaced pre-existing European stocks. Although the leading edge of the invasion appeared to be dominated by African matrilines [16], there was also mixing and phenotypic expression of nuclear genes [17] due to drones of each race mating with queens of the other. Notably, a substantial contribution (20-30%) of European nuclear genes remains in the Africanized population [18,19]. In the long run, although a mixture of various African matrilines eventually dominate in the population, some European matrilines persist (with the M lineage being much more successful in the tropics than the C (Italian/Carniolan) lineage) [20].
Practical answer: Our current state of knowledge does not allow me to definitively rank the relative importance of matrilines and drone contributions to successive generations. Each appear to play an important part in natural selection, with certain matrilines predominating under certain environmental circumstances.
Practical note: Pinto [21] points out that the strong selective pressure by varroa may have been a critical contributing factor in the introgression of the African genome into the Texas bee population, since the mite effectively wiped out the European feral competition, opening up an unexploited niche to the “Africans.” The point is that no matter how “fit” a certain breed may be at the moment, the introduction of a single parasite or other novel environmental factor may immediately make previously successful genetic combinations obsolete.
Another thought: keep in mind that in Mediterranean and temperate Europe, the C and M lines held their own against the African matrilines (from which they originally descended). It could well be that in the U.S., once the European stocks evolve resistance to varroa, that they may reclaim territory currently held by the Africanized bees.
We still have a great deal to learn about breeding honey bees for optimum performance and survival. I plan to return to this subject later on in this series when I discuss the future of bee breeding.
The Grass Is Always Greener
Speaking of breeding, it seems to be human nature to tend to think that bees from somewhere else would be better than the local stock. In actuality, it depends upon what one means by “better.” For early beekeepers, “better” likely meant less stinging, and perhaps a propensity to store a greater amount of honey. Some 3000 years ago commercial beekeepers in Israel imported a more gentle and productive race of bee from Turkey [22]. For similar reasons, beekeepers in this country came to prefer gentle Italian, Carniolan, and Caucasian stocks (all C lineage) over the originally imported (and successfully invasive) “German dark” bees (M line).
And for commercial migratory beekeepers today, Italian- or Carniolan-type stocks are justifiably preferred, just as White Leghorn chickens or Angus cattle are preferred by their respective producers.
Practical application: migratory beekeepers need high-performance bees specifically adapted to the commercial niche, which includes early buildup, intense crowding, rough handling, heavy supplemental feeding, regular treatments for parasites, and exposure to ag chemicals and miticide residues. The U.S. queen market predominately caters to these beekeepers, thus, breeders select for appropriate traits. Nothing in this article should be taken as a criticism of commercial stocks.
Locally-Adapted Stock
But as we could see from the pie charts in my previous article, Nature may exert a far different selective pressure than do commercial bee breeders. Nature selects for bees that survive without human help in their specific environment. Such bees are termed “locally adapted.”
Note that the concept of geographic “local adaptation” is difficult to apply to migratory bees that may be moved from Texas to California to Washington to North Dakota all in one season. But how about stocks for stationary beekeepers who wish to maximize their profits by minimizing the costs of feeding and treatments? Could they benefit from keeping bee stocks that are better adapted to local conditions [23] (see Fig. 3, in which I continue to outdo myself with colorful graphics)?
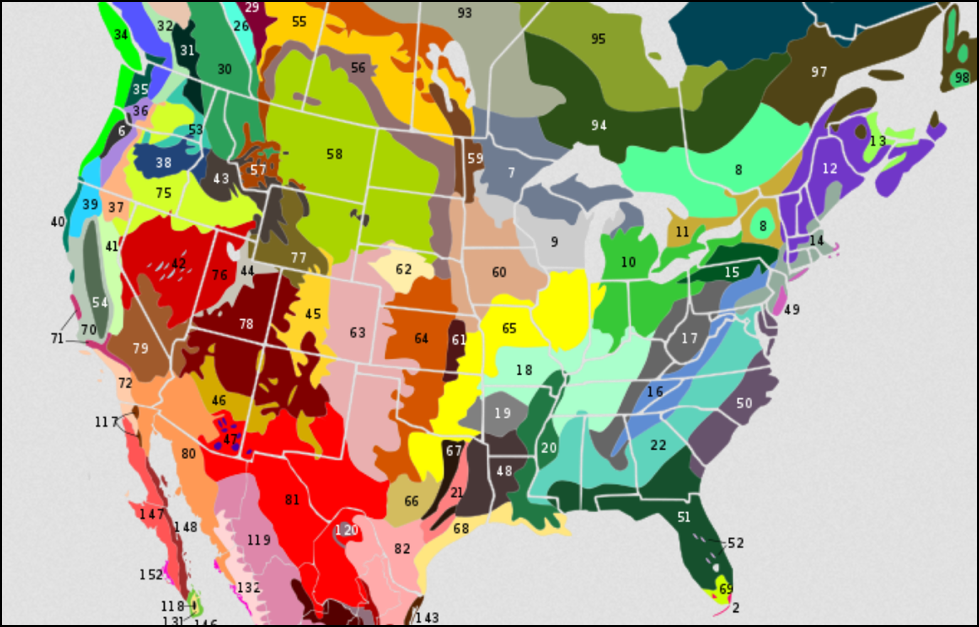 Figure 3. In which ecoregion do you keep bees? There is absolutely no reason to expect that bees selected for fitness in any one of the differently-colored ecoregions above would perform well in another region. Source: TH Ricketts [[i]].
Figure 3. In which ecoregion do you keep bees? There is absolutely no reason to expect that bees selected for fitness in any one of the differently-colored ecoregions above would perform well in another region. Source: TH Ricketts [[i]].
[i] Ricketts, TH, et al (1999) Terrestrial ecoregions of North America : a conservation assessment. Island Press. From Wikimedia. The key to the name of each numbered ecoregion can be found at http://commons.wikimedia.org/wiki/File:Terrestrial_ecoregions_USA_CAN_MEX.svg. I personally breed for bees adapted to a small portion of Region 41.
{kind=link}
As Büchler [25] points out:
In the majority of selection and breeding programs, economic traits (such as honey productivity and colony strength) together with traits desirable for modern beekeeping (such as gentle temper and low swarming tendency) have been of predominant importance. In contrast, disease resistance, viability, and adaptation to local conditions were considered less important, as deficiencies in these characters could often be compensated by pharmaceuticals, artificial feeding, and other management techniques.
Practical question: the honey bee as a species exhibits extreme plasticity (as evidenced by the numerous indigenous races), and can be bred for nearly any combinations of traits. The question to me, in this day of 30% winter losses despite the time and money invested in feeding, medicating, and mite treatments, is whether we’ve been breeding for the right traits?
When we breed stock for the benefit of high production, it typically comes at a cost in overall fitness. As Charles Darwin observed in 1857, “…as Goethe expressed it, ‘in order to spend on one side, nature is forced to economise on the other side.’” Evolutionary ecologists term this relationship “tradeoffs in fitness components.” With the level of colony mortality that many beekeepers are experiencing with today’s bee breeds, we may wish to reevaluate the cost/benefit ratio of the traits that we select for (Fig. 4).
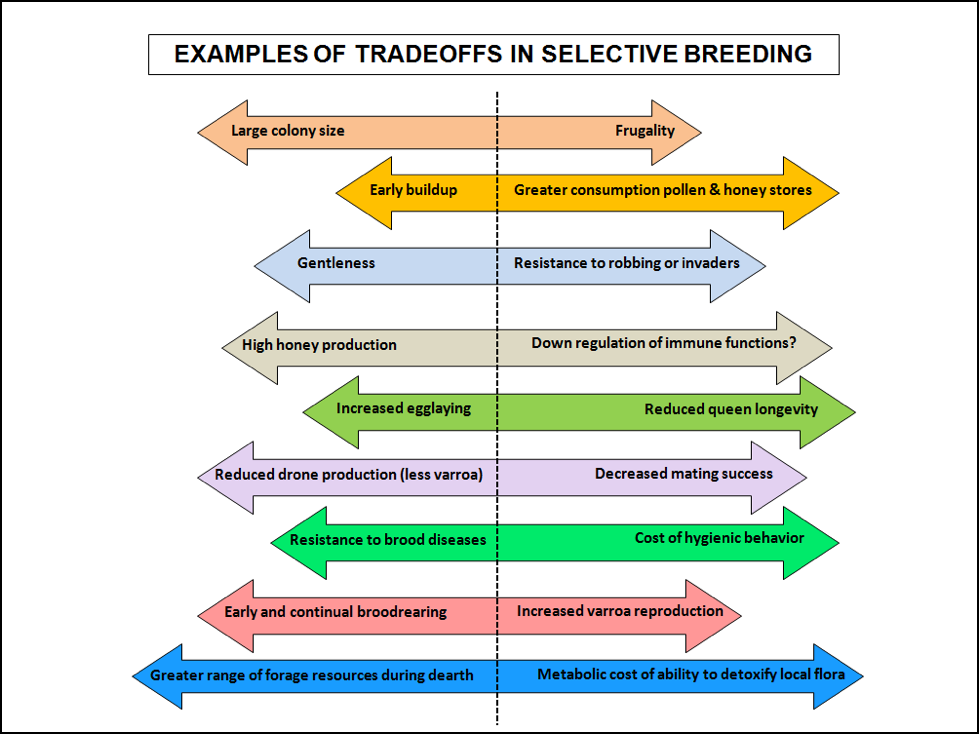 Figure 4. There are always tradeoffs in the breeding of bees. When you select your breeding stock, you must think about the direction you wish to move each arrow—every trait that you select for comes at a cost.
Figure 4. There are always tradeoffs in the breeding of bees. When you select your breeding stock, you must think about the direction you wish to move each arrow—every trait that you select for comes at a cost.
To a biologist, it just seems like common sense that a bee stock adapted to the local environment would be expected to survive better with minimal management than would an exotic breed. Think about it—the process of natural selection would have already done most of the work for us. Bayer [26], speaking of livestock in general, explains:
Generations of natural and deliberate selection have resulted in local breeds with a high degree of disease resistance or tolerance and capable of subsisting on the available feed resources, which are seasonally scarce and of low quality.
The breeders of other species of livestock are showing great interest in the breeding potential of locally-adapted breeds, especially for parasite resistance and ability to survive in inhospitable areas [27]. As forhoney bees, a recent issue of the Journal of Apicultural Research is devoted to this very subject [28]. Let’s see what the researchers found:
Francis [29] concluded that:
Our results suggest that bees of local origin are better in tune with environmental factors related to flowering patterns, climatic variation and locally prevailing apicultural methods and therefore may command more sufficient resources to fend off pathogens.
Hatjina [30] found that:
The local genotypes showed a trend to collect more honey than the non-local ones, which shows their ability to develop higher adult bee populations…and better ability to forage on the local flora. This adaptation and their longer survivorship…could also explain the fact that the survived colonies (most of them of local genotypes) had higher honey production during the second year.
Costa [31] in Italy found that locally-adapted Italian stocks produced more honey than Italian stocks imported from other parts of the country. Dražić [32], testing Carniolan stocks in Central Europe found that:
At the end of test period, colonies of introduced genotype had lower survival rates at both locations.
Buchler [33] explains:
Ecological studies have shown that genotype × environment interaction is an indicator for local adaptation and fitness. The most important form of this interaction is antagonistic pleiotropy [34], whereby different alleles have opposite effects on fitness in different habitats. This implies that no single genotype is superior in all environments, leading to a trade-off in adaptation to different habitats…
Most of the theory relevant for local adaptation concentrates on the evolution of ecological specialization, assuming a trade-off in fitness across habitats mediated by a quantitative trait or traits… In simple words, local adaptation should result in improved fitness of each population in its own habitat…
In our study, we observed a significantly higher survivorship of the local genotypes compared to the non-local ones, clearly indicating a specific local adaptation of the honey bee populations considered in the experiment.
Furthermore, the results of our study show that it is not merely an ecological issue, but also a commercial one: the use of local honey bee populations provides a higher chance of colony survival, and the use of maladapted bees attributes to high colony losses, as recently observed in many regions. Thus, local breeding activities should be promoted and encouraged throughout the native range of Apis mellifera.
Wrap-Up
What with all the health issues that we’re having with bees these days, we may want to take a closer look at “survivor stocks” that manage to stay alive without human assistance. I suspect that our feral stocks are an invaluable and underutilized resource. The challenge will be to use them as base stocks from which to select those that are workable and productive, without losing their toughness [35]. The first step will be to start propagating stocks naturally adapted to each of the colored ecoregions in Figure 3.
Practical application: what are you waiting for? It is unreasonable to expect the large queen producers in the West and South to breed for bees specifically adapted to your area. I have asked for shows of hands from audiences all over the continent, and can assure you that there is a strong demand for locally-adapted mite-resistant stock. Entrepreneurial beekeepers could produce mid- to late-season queens for regional sale, and they would be a bargain at $30 each to local beekeepers. This is a potential business opportunity for those wishing to expand their income stream.
Next
O.K., I got sidetracked. I hope to return to the effects of early bee breeding on the realized niche of the honey bee, and how it set the stage for the devastation by varroa.
Acknowledgements
Thanks as always to my long-time collaborator Peter Loring Borst. And especially to those researchers who have done, or are doing, the hard work involved in answering our questions about bee genetics, fitness, and selective breeding.
Footnotes And Citations
[1] Or perhaps not. Some of the ancestral races consists in only a tiny remnant patches http://en.wikipedia.org/wiki/Zea_%28genus%29. Note that despite the fact that although maize is widely planted across the world, I can find no reference to it ever existing in an escaped form. Two interesting and well-illustrated reads on the breeding of corn can be found at http://www.businessinsider.com/the-story-behind-glass-gem-corn-2013-10?op=1 and by Googling “The Ancestry of Corn” by George Beadle.
[2] The benzoxazinoid DIMBOA, a natural insecticide produced by ancestral maize. http://www.wzw.tum.de/index.php?id=185&L=1&tx_ttnews[tt_news]=44
[3] Maize breeders have stored over 135,000 varieties of maize germplasm in seed banks, and draw from the ancestral teosinte species for desirable traits.
[4] With the caveat that some genotypes only work well with some mitotypes.
[5] Kraus, FB, et al (2007) Asymmetric introgression of African genes in honeybee populations (Apis mellifera L.) in Central Mexico. Heredity (2007) 99, 233–240. Open access.
[6] Actually, various alleles of those genes. In this diagram, I arbitrarily used 8 dots to represent the degree of potential allelic diversity.
[7] In reality, a queen only passes half of her nuclear alleles to an egg, so individual queens would not pass all the alleles shown.
[8] In bee breeding, we use the loosest definition of “hybrid”—intraspecific hybridization between races or breeds of the same species. Intraspecific hybrids are generally fertile, although some may exhibit a lack of fitness, depending upon how closely related the races were.
[9] Pinto, M A, et al (2005) Africanization in the United States: replacement of feral european honeybees (Apis mellifera L.) by an African hybrid swarm. Genetics 170:1653–1165. Open access. This is a great review of the genetic process of Africanization in Texas.
[10] Jensen, AB, et al (2005) Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Molecular Ecology 14: 93– 106.
Quezada-Euán, JJG, et al (2003) Hybridization between European and African-derived honeybee populations (Apis mellifera) at different altitudes in Perú. Apidologie 34: 217–225.
[11] O.K., if you’ve got your reading glasses on and look at wing veination you might be able to.
[12] http://perso.fundp.ac.be/~jvandyck/homage/elver/
[13] Koeniger, G (2005) The neglected gender – males in bees. Apidologie 36: 143–144.. http://www.apidologie.org/articles/apido/pdf/2005/02/M36200f.pdf
[14] I am certainly no authority on this subject, and would appreciate knowledgeable feedback if I’ve gotten something wrong.
[15] Remember that the haploid drones carry only one copy of each gene, whereas the diploid queens and workers carry two copies, and potentially two different alleles.
[16] Muralidharan, K and HG Hall (1989) Evidence from mitochondrial DNA that African honey bees spread as continuous maternal lineages. Nature 339: 211 – 213. Open access.
[17] Degrandi-Hoffman, G, et al (2003) Patriline composition of worker populations in honeybee (Apis mellifera) colonies headed by queens inseminated with semen from African and European drones. Apidologie 34: 111–120. Open access.
[18] Well reviewed in the following papers:
Quezada-Euan, JJG (2000) Hybridization between European and Africanized honeybees in tropical Yucatan, Mexico. II. Morphometric, allozymic and mitochondrial DNA variability in feral colonies. Apidologie 31: 443–453. Open access.
Nilza Maria Diniz, NM, et al (2003) Genetic structure of honeybee populations from southern Brazil and Uruguay. Genet. Mol. Biol 26 (1). Open access.
An interesting discussion on the “gentle” AHBs in Puerto Rico: Rivera-Marchand, B, et al (2012) Gentle Africanized bees on an oceanic island. Evolutionary Applications 5: 746–756. Open access
[19] Szalanski, AL and RM Magnus (2010) Mitochondrial DNA characterization of Africanized honey bee (Apis mellifera L.) populations from the USA. Journal of Apicultural Research and Bee World 49(2): 177-185. Open access.
[20] Pinto, op cit.
[21] Pinto, op cit.
[22] Bloch, G, et al () Industrial apiculture in the Jordan valley during Biblical times with Anatolian honeybees. PNAS 107(25): 11240–11244. http://www.pnas.org/content/107/25/11240.full
[23] For a fascinating read on local adaptation by grazing animals (with a look to the future, and in many ways applicable to bees), see Provenza, FD (2008) What does it mean to be locally adapted and who cares anyway? J. Anim. Sci. 2008. 86(E. Suppl.):E271–E284. http://www.journalofanimalscience.org/content/86/14_suppl/E271.full.pdf+html Something that I found of great interest was local adaptation to toxic plants. Locally-adapted stocks can utilize a greater range of plant forage species during times of dearth, due to their ability to deal with plant allelochemicals. See https://scientificbeekeeping.com/sick-bees-part-18f2-colony-collapse-revisited-plant-allelochemicals/
[24] Ricketts, TH, et al (1999) Terrestrial ecoregions of North America : a conservation assessment. Island Press. From Wikimedia. The key to the name of each numbered ecoregion can be found at http://commons.wikimedia.org/wiki/File:Terrestrial_ecoregions_USA_CAN_MEX.svg. I personally breed for bees adapted to a small portion of Region 41.
[25] Büchler, R, et al (2010) Breeding for resistance to Varroa destructor in Europe. Apidologie 41: 393–408. Open access.
[26] Bayer, W (1989) Low-demand animals for low-input systems. ILEIA Newsletter 5(4). Open access.
[27] Hanotte, O, et al (2010) Time to tap Africa’s livestock genomes. Science 328: 1640–641. Open access.
Lamy, L, et al (2012) Factors Influencing Livestock Productivity. In V. Sejian et al. (eds.), Environmental Stress and Amelioration in Livestock Production. Springer-Verlag. Open access. This paper goes into considerable detail as to the sorts of specific adaptations livestock can make to local environmental parameters.
[28] Most of the immediately following studies are in a special issue of the Journal of Apicultural Research, and are open access.
[29] Francis, RM, et al (2014) Effect of genotype and environment on parasite and pathogen levels in one apiary – a case study. Journal of Apicultural Research 53(2): 230-232.
[30] Hatjina, F, et al (2014) Population dynamics of European honey bee genotypes under different environmental conditions. Journal of Apicultural Research 53(2): 233-247.
[31] Costa, C, et al (2012) Differences in colony phenotypes across different origins and locations: evidence for 241 genotype by environment interactions in the Italian honey bee (Apis mellifera ligustica). Apidologie 43(6): 634-642.
[32] Dražić, MM, et al (2014) Colony development of two Carniolan genotypes (Apis mellifera carnica) in relation to environment. Journal of Apicultural Research 53(2): 261-268.
[33] BÜCHLER, R, et. al.(2014) The influence of genetic origin and its interaction with environmental effects on the survival of Apis mellifera L. colonies in Europe. Journal of Apicultural Research 53(2): 205-214.
[34] A gene that is pleiotropic affects more than one trait. In antagonistic pleiotropy, there can be a trade-off between the beneficial and the detrimental effects of a single gene. Lobo, I (2008) Pleiotropy: One gene can affect multiple traits. Nature Education 1(1):10. http://www.nature.com/scitable/topicpage/pleiotropy-one-gene-can-affect-multiple-traits-569
[35] This challenge is most evident in areas that are fully Africanized, such as Texas, Arizona, and Southern California, and I commend breeder Danny Weaver on his efforts. Compared to breeding from Africanized stock, selection from indigenous feral European stocks would be expected to be far easier.