
A Study on Bee Drift and Mite Immigration: Part 1

May 8, 2023
Was Another Field Study Justified?
The Biology Behind Mite Transfer Between Hives
How Much Mite Immigration Actually Occurs?
How Do the Mites Get Transferred?
How Important is the Drifting of Drones?
How Much Drift is There of Worker Bees?
How Important are the Guard Bees?
Do Mites and Viruses Cause Increased Drifting?
Robbing vs. “Ordinary” Drifting
How Far Can Mites Hitchike a Ride?
A Study on Bee Drift and Mite Immigration
Part 1
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ in February 2023
Many beekeepers complain about sudden late-summer spikes in their varroa counts, blaming them on mites immigrating into their hives from other colonies in the neighborhood. When my sons and I suspected that this was taking place in some our own yards, I decided to run a field study to find out exactly what was happening.
INTRODUCTION
It requires human transport to move the varroa mite across geographical barriers, but to the dismay of beekeepers, once introduced into a country, the mite can spread rapidly to all colonies in the region. This invasiveness is largely due to the natural drift of bees, which also results in the carrying of mites from hive to hive. I wrote about what was then known about mite drift back in the spring of 2018, and suggest that if you’re interested in this subject that you first review my two articles on the subject [[1]].
SO WAS ANOTHER FIELD STUDY ON MITE DRIFT JUSTIFIED?
To answer that question, let’s take a look at data from Ritter in Germany, taken back in 1987 [[2]]. The paper was in German, but worth translating. Ritter set up two separate apiaries of 16 mite-free colonies — one in the foothills of the Black Forest in an area with low colony density and the other in the Rhine plain with numerous heavily infested colonies in the environment, and then tracked the infestation rates of the bees and brood in the experimental hives over time. His results confirmed that having heavily-infested colonies in the neighborhood can make a heckuva difference (Figure 1).
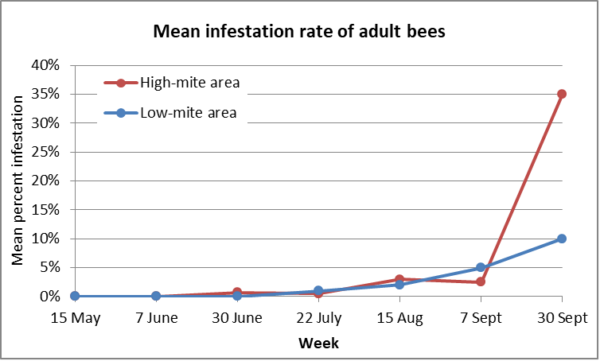
Fig. 1. Of interest is how long it took for the effect to show in the mite washes. At an infestation rate of 35%, that would be 110 mites in a wash of a half cup of bees! Side note: Back then the recommendation in Germany was to use gasoline for performing mite washes. I assume that they were careful with their smokers!
But was the above sudden spike in the infestation rate due to a sudden influx of mites in September? Take a look at Figure 2.
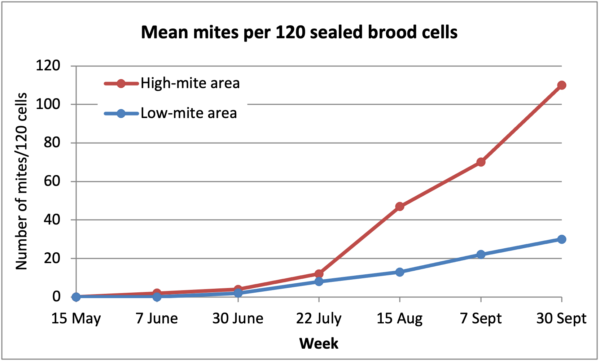
Fig. 2. It appears that the impact of mite immigration actually started to take place long before September. The apparent “sudden” spike as indicated by mite washes may have actually been due to an exponential increase in mite emergence in September, into a decreasing number of adult bees in the hives. This may explain why even beekeepers who are monitoring via mite washes may still get surprised!
Practical application: Ritter’s field data certainly suggests that mite immigration from other colonies in the neighborhood can affect the rate of mite buildup in one’s own hives. So I wanted to dive deeper.
In order to better understand this phenomenon, I modified my mite model [[3]] to run some simulations (after first confirming that the model could produce results in the ballpark of Ritter’s data). Let’s first take a look at the estimations of details of varroa buildup over time in an untreated colony headed towards collapse in October (Figure 3).
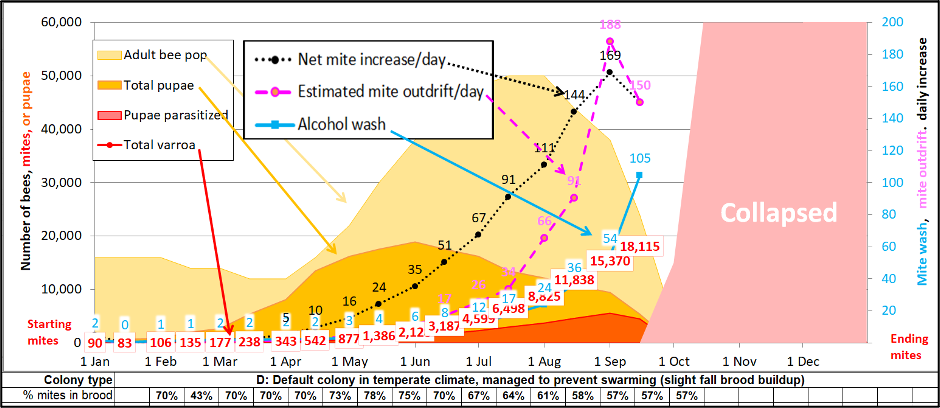
Fig. 3. Allow me to use the simulation above to introduce you to the concept of the “Mite Factory.” I added extra plots for daily amount of mite increase, as well as for the estimated amount of mite exodus on outgoing bees. Read below for explanations.
By the time an untreated, nonresistant Mite Factory about to collapse:
- It will typically contain around 15,000 mites in total.
- The above number will be increasing at a net increase of about 150 mites per day.
- And due to the short lifespan of summer bees, above that number, there will likely be over 150 mites hitching a final ride out of the colony each day [[4]].
Unfortunately, there are too many variables for the model to predict the number of those exiting mites lucky enough to be carried to another hive. With the above figures in mind, you may be wondering:
Practical question: What impact might a Varroa Factory in the neighborhood have upon my well-managed (treated) hives?
To answer that question, let’s run the same simulation, but this time with a couple of efficacious treatments (Figure 4).
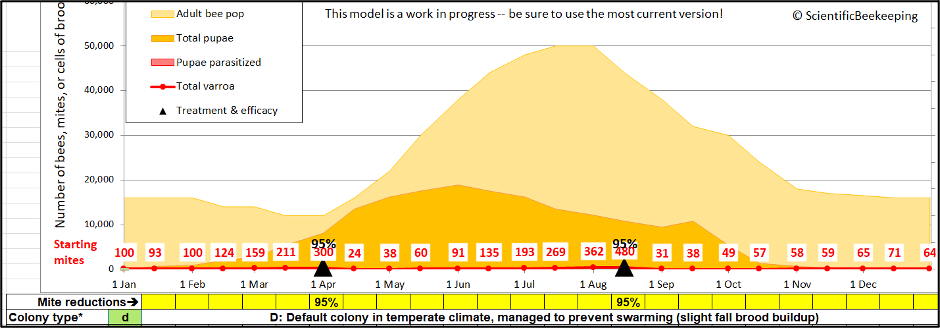
Fig. 4. Same simulation, but this time treated in April and August, resulting in a far lower mite level in mid-September (around 35). This now gives us two baseline numbers (15,000 and 35) to play with.
Now imagine a neighborhood in which one out of every ten hives is untreated, while the other nine colonies are treated (each containing an average of only 35 mites, as in the second simulation). Let’s see what happens if an arbitrary 10% of the 15,000 mites in the untreated colony somehow manage to evenly “diffuse” (via bee drift) into the nine other treated hives (an average of 165 mites per hive)(Figure 5).

Fig. 5. A single “Mite Factory” has the potential to seriously reinfest all the other hives in the neighborhood after their mid-August treatment (in reality, the distribution of mites would of course be uneven, with some colonies receiving more mites than others). Imagine the effect if your apiary was surrounded by a number of “mite factories”!
OK, so mite immigration could clearly have a noticeable impact if you already had your mites well under control, but what if you were running a little behind the curve? (Figure 6).
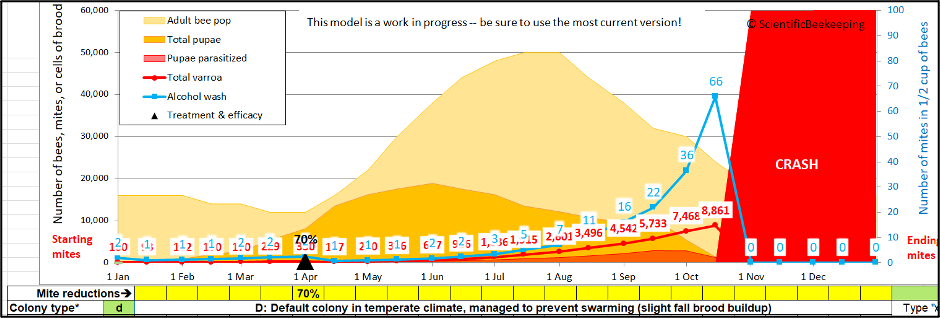
Fig.6. In this simulation of fairly-typical colony management, the efficacy of the April treatment was only so-so (due to most of the mites being in the brood). And then the beekeeper hadn’t gotten around to giving an August treatment. Going into September, the simulated colony shows a mite wash count of 16 — which would be typical of “average” management practices [[5]]. But the mite population in that hive would soon exceed 5000, meaning that an immigration of the same 165 mites as in the previous scenario would be only a drop in the bucket!
Practical application: If you’re not staying on top of varroa all season long, it may be a stretch to blame a late-season spike in mite counts upon mite immigration from elsewhere.
It appears as though mite immigration could indeed be an issue for those making the effort to maintain low mite counts. So I felt that a field study to better understand mite immigration in my operation was justified!
BACKGROUND RESEARCH
The first thing I want to do when designing a research project is to find out what is already known. For this article, I’ll snip some quotes (with minor paraphrasing and added emphasis) from relevant publications.
The second thing is then determine the specific questions that I want to try to answer — these will be my study objectives (which I will list below).
THE BIOLOGY BEHIND MITE TRANSFER BETWEEN HIVES
Varroa is a very successful parasite, rapidly reproducing in its own host honey bee colony. But for the parasite’s continued success, bloodlines of the mite must somehow “horizontally transfer” to other host colonies prior to the death of their host colony.
To the mites’ great advantage, they’re able to utilize bees exiting their host colony as an unwitting delivery service that takes them directly into other colonies within flight range. This hitchhiking not only allows varroa to infest newly-established colonies, but also to mix their genes among neighboring host colonies (which helps the mite to evolve).
HOW MUCH MITE IMMIGRATION ACTUALLY OCCURS?
It takes only a single fertile female mite to start an infestation in a colony. But beekeepers soon noticed that the degree of mite immigration was sometimes much more than that. Following Ritter’s publication, in 1988, German researchers Sakofski and Koeniger wrote a chapter called “Natural transfer of Varroa jacobsoni among honeybee colonies in autumn” [[6]]. And in 1990 [[7]] they again confirmed that colonies from which varroa had been eliminated by treatment could be reinvaded by the mite, especially during late summer and fall. And then in 1992Greatti [[8]] published data indicating stunning amounts of mite invasion taking place in his monitored colonies in Italy. These findings were followed by other research, notably that of Frey [[9]], who found that mites could drift for a considerable distance. I’ve summarize the above study results previously [[10]], but would they apply to my locations in California?
Practical application: There’s no doubt that mite migration between colonies can take place, and that it can be substantial. The questions for beekeepers then are exactly how and why it occurs, how much immigration a colony can actually receive, and is there anything that we can do to reduce it.
Study Objective #1: To determine the amount of mite immigration that actually takes place in late summer and fall where I keep my bees.
Study Objective #2: To determine whether that mite immigration is steady or episodic.
Study Objective #3: To determine whether all hives suffer equally from mite immigration, or whether some are more attractive or receptive than others.
HOW DO THE MITES GET TRANSFERRED?
Since the mites depend upon bees to carry them to other colonies, it’s important to understand this aspect. Greatti [[11]] listed the three main methods by which the mite manages to disperse from one colony to another:
- By the drifting of drones, which are known to drift freely between hives.
- By the drifting of worker bees — which will tend to “diffuse” mites from colonies with high infestation to those with low infestation.
- By the robbing of colonies weakened by heavy infestation.
HOW IMPORTANT IS THE DRIFTING OF DRONES?
A recent study by Mortensen [[12]] confirmed that drones do carry mites. It’s well known that drones tend to drift to other hives. That said, Greatti, along with quantifying the actual amount of mite immigration that could take place, tested whether placing drone excluders over the entrances of colonies would reduce immigration. He found that it did not.
Practical application: Although drones do a lot of drifting, they may not contribute much to mite drift, and since there are very few drones in our colonies in late summer, I didn’t pursue the drone factor.
HOW MUCH DRIFT IS THERE OF WORKER BEES?
There have been plenty of studies indicating that there can be considerable drift of worker bees, especially in crowded apiaries, where more than half the workers in a colony may drift at some time in their lives to other hives [[13]].
But how about the relatively sudden disappearance of the bees from a hive as it collapses from the varroa-virus complex. Do those bees just “disappear,” or do they wind up in other colonies?
Study Objective #4: To determine the proportion of workers that drift from collapsing colonies into other hives.
HOW IMPORTANT ARE THE GUARD BEES?
There’s another aspect of worker drift besides bees just getting lost — to enter a foreign hive, they first need to get past the guards. Since I have long selected for “gentle” stock, I’ve wondered whether I’ve also been inadvertently selecting for bloodlines whose guards might be more permissive to drifted bees attempting to enter their hive (Figure 7).

Fig. 7. Although we breed for gentleness, I often observe guards doing a good job at defending the entrance against intruders.
In 2017 Bordier [[14]] used video cameras to track tagged bees at the entrances of monitored hives (Figure 8).

Fig. 8. I visited the INRA research facility in 2018, and snapped this picture of one of the video entrance monitoring units that they were using.
Bordier concluded that (I’ve boldfaced the highlights):
Drifting behavior is a consequence of orientation errors, and also depends on the acceptance level of the new host colony… changes in drifting intensity are likely the result of an increase in colony acceptance. In normal conditions, only 3.5 % of incoming bees are checked by guard bees, but this permissiveness changes according to season, density of bees and nectar and pollen resources— the acceptance levels of colonies increase when nectar flow is abundant but decrease during dearth periods.
Practical application: So I’m still not clear on the importance of the guards in preventing mite immigration. Of interest is the finding by Cappa [[15]] that guard bees mount a greater defense against drifting bees that have been parasitized by a mite (how much might this play into “mite resistance” of a colony?). Contrast that finding to that of Geffre [[16]], who “found that IAPV-inoculated bees were significantly more likely to be accepted by the guards at the colony entrance than were…controls.”
We obviously still have much to learn about guarding and drifting. I set up my study so that we would only recover drifted bees that made it past the guards on the landing board.
DO MITES AND VIRUSES CAUSE INCREASED DRIFTING?
It is well known to biologists that any number of parasites can change the behaviors of their hosts in order to effect better transmission [[17]]. So as proposed by DeGrandi-Hoffman [[18]] in 2017:
We hypothesize that stress from parasitism and virus infection combined with effects that viruses have on cognitive function may contribute to forager drift and mite and virus dispersal.
So is there evidence for increased drift from colonies undergoing collapse? In 1991 Sakofski [[19]] found that:
By comparing numbers of bees drifting from highly infested and noninfested small colonies it could be shown that infested colonies drifted more frequently. The invasion of mites into bee colonies was monitored during a period from spring to autumn. All colonies showed low values of mite influx which increased considerably at the end of July and remained high until the beginning of October. Individual values differed over wide ranges which are likely to be caused by robbery.
But in 2000, Neuman [[20]], although finding substantial drift of workers and drones, did not find a correlation between worker bee drift and varroa infestation
Goodwin [[21]] ran a field trial in New Zealand wrote in 2006, in which six of eight high-mite colonies with marked bees collapsed and got robbed out. Despite that:
The average percentage of bees drifting from untreated hives infested with varroa into neighboring treated hives did not exceed 3%. There were no significant differences between the percentage of drifting bees from the varroa-infested colonies and the treated colonies at any stage throughout the trial. This suggests that the reinfestation of treated colonies does not predominantly result from heavy varroa infestations causing worker bees to drift to other colonies, more than they generally do in practically varroa-free colonies.
And then in 2014 the premise that highly-infested bees were predisposed to drift was given wind by Cervo [[22]]:
At low mite abundance, mites stay within the colony where they are born and promote their reproduction by riding nurses. This preference ensures that mites are quickly transferred to another host larva within the same colony where they can reproduce. When mite abundance increases within the colony, the lack of differences in chemical cues between nurses and foragers probably does not allow mites to discriminate between bees with different tasks and causes mites to ride on both of them…. by riding both nestmate foragers and foreign [robbers], mites increase their probability of leaving the exploited colonies. An infested colony, progressively weakened by parasites and mite-transmitted diseases, finally becomes an easy target for robbing foragers from foreign colonies; these robber foreign bees might represent excellent vectors to directly transfer mites from the old exploited beehive to a new one.
This hypothesis immediately gained traction, and by 2017 the term “Mite Bombs” was being thrown about, although with little supportive evidence. And it was a foggy notion – not clearly differentiating the day-by-day “diffusive” mite drift from high-mite or collapsing colonies, versus the sudden and short-term direct transfer of mites during robbing events that may occur during the collapse of a colony.
In 2017 Bordier [[23]] found that:
Approximately 10 % of our tracked bees drifted into a foreign colony. The drifting prevalence was influenced by the colony’s location in space but not by N. ceranae parasitism.
There were no significant differences between the percentage of drifting bees from the varroa-infested colonies and the control colonies at any stage throughout the study. This suggests that the reinfestation of treated colonies does not predominantly result from heavy varroa infestations causing worker bees to drift to other colonies, more than they generally do in practically varroa-free colonies….The normal level of drift in apiaries will, however, spread varroa from untreated to treated colonies.
And in 2019 Peck and Seeley [[24]] found that:
We saw no indications that drifting of bees from the mite donor colonies into the mite receiver colonies increased as the mite levels in the mite donor colonies increased.
Practical application: As attractive as a virus-induced increase in drifting behavior hypothesis is, there are to date conflicting findings (Figure 9).

Fig. 9. It’s easy to identify an unfortunate worker that suffered from such a high infection rate of DWV that its wings could not develop. But you can safely assume that every worker in this photo is infected with DWV to some extent, as evidenced by the dying brood. But does the varroa-DWV complex cause these infected bees to drift at a higher rate?
Study Objective #5: To determine whether elevated varroa/virus levels increase bee drift in my own operation.
ROBBING VS. “ORDINARY” DRIFTING
We’ve now worked our way down to whether mite immigration (a) is primarily due to the “ordinary” drift of bees from hive to hive (which I call “mite diffusion” since the net effect will be to move mites from colonies with high mite concentration to those with lower concentration), or (b) is mainly from robbing bees carrying mites back from varroa-weakened hives. Such robbing could be surreptitious (and unnoticed by the beekeeper) or “overt” (easily observed at the entrance when a collapsing colony gets mobbed by robbers) (Figure 10).

Fig. 10. The unusual activity at the entrance, coupled with the debris on the landing board indicates that this dwindled colony is getting robbed. I suspect that in our location, most mite immigration due to robbing might be from the surreptitious robbing that takes place during nectar flows, when there are few guards at the entrance.
Peck and Seeley [[25]] looked into mite drift during robbing events taking place in Ithaca, NY late in the season:
Our data also reveal that the colloquial terminology for [heavily infested] colonies—“mite bombs”—does not accurately describe the mechanisms of intercolony mite transmission that we observed in this study. We saw no sudden “explosion” of mite-carrying bees from sick colonies to each of the healthy colonies via worker drifting. Indeed, we found that few mites passed from the heavily infested (MDC) colonies to the weakly infested (MRC) colonies through either worker drift (mostly to nearby colonies) or drone drift. It was only when the MDCs were weakened so much that they became irresistible robbing targets that mites passed in large numbers from the MDCs to the MRCs.
Practical application: The mites in a colony that is being robbed as it is undergoing collapse (while there are still plenty of live mites in the hive) can certainly hop onto the robbers (which the mites could easily recognize due to their different odor) and get carried back to the robbing colony. But such immediate robbing doesn’t take place in all landscapes — see my ancillary article in this issue.
And finally, Kulhanek published a study last year [[26]].
More bees from low mite colonies (n= 37) were detected in receiver apiaries than bees from high mite colonies (n= 10). Receiver colony Varroa population growth was associated with visitation by non-natal bees, but not high mite bees alone. Finally, colonies lacking robbing screens experienced faster Varroa population growth than screened neighbors. Results indicate visiting non-natal bees may vector mites to receiver colonies. These results do not support the current two leading theories regarding mite immigration – the “mite bomb” theory (bees from high mite colonies emigrating to collapsing colonies), or the “robbing” theory (natal robbing bees return home with mites from collapsing colonies).
Practical application: Although mite drift during rob outs can indeed take place, it may be that mite diffusion due to ordinary bee drift may be more important. Since robbing screens don’t prevent colonies from robbing, Kulhanek’s findings suggest that they may reduce bee (and therefore, mite) drift (teaser: I will soon be publishing my own testing of robbing screens).
Study Objective #6: To determine whether mite immigration correlates with robbing.
There’s still one more item of interest…
HOW FAR CAN MITES HITCHIKE A RIDE?
A study by Frey in 2011 [[27]] had a surprising finding — that mite drift is not necessarily a function of distance, and that there can be considerable drift even to colonies a mile distant.
During the experimental period, between 85 and 444 mites per colony were introduced into the receiver colonies. There were no significant differences in the invasion rates in relation to the distance between donor and receiver colonies. In total, 2,029 mites were found in the 10 receiver colonies, but these only correspond to 2.5% of the total mite population in the donor colonies at the start of the experiment. This means that the major part of the initial V. destructor population died together with the collapsed host colonies…From a practical perspective we could show that highly infested honey bee colonies present a substantial risk to already treated colonies up to distances of 1.5 km [roughly a mile] away.
Kulhanek [[28]] observed substantial drift between hives in the same yard, but her cameras observed only 47 marked bees (out of approximately 32,000 painted) drifting to receiver hives a half mile or further distant.
Practical application: Frey’s finding of the lack of correlation between mite drift and distance was something that I wanted to test in my home yard.
Study Objective #7: To determine the distance that bees can drift from collapsing colonies.
Study Objective #8: To determine how the rate of drift to nearby hives compares to that to hives more distant.
NEXT MONTH
In the next installment I’ll show we set up our field study in order to find out the answers to the listed objectives. Stay tuned!
ACKNOWLEGEMENTS
Thanks to Peter Borst and Dianne Behnke for their assistance with finding and scanning copies of old research papers.
CITATIONS AND NOTES
[1] The Varroa Problem: Bee Drift and Mite Dispersal, Parts 16 a & b:
https://scientificbeekeeping.com/the-varroa-problem-part-16a/
https://scientificbeekeeping.com/the-varroa-problem-part-16b/
[2] Ritter, W., & E Leclercq (1987). Entwicklung der Bienen-und Varroa-population in Gebieten mit unterschiedlichen Möglichkeiten der Reinvasion. Tierärzt Umschau 42: 548-51.
[3] https://scientificbeekeeping.com/scibeeimages/Randys-Varroa-Model-V2022-1-1.xlsx
[4] This figure is calculated from the calculated number of non-returning adult bees per day (due to aging, getting lost, or other causes), assuming that they are carrying mites at only 2/3 the infestation rate of the worker bees on average).
[5] Kulhanek K, et al (2021) Survey-derived best management practices for backyard beekeepers improve colony health and reduce mortality. PLoS ONE 16(1): e0245490.
[6] Sakofski, F & N Koeniger (1988) Natural transfer of Varroa jacobsoni among honeybee colonies in autumn. In European Research on Varroatosis Control.
[7] Sakofski, F, N Koeniger, S Fuchs (1990). Seasonality of honey bee colony invasion by Varroa jacobsoniOud. Apidologie, 21(6), 547-550.
[8] Greatti, M, N Milani, F Nazzi (1992). Reinfestation of an acaricide-treated apiary by Varroa jacobsoniOud. Experimental & applied acarology, 16(4), 279-286.
[9] Frey, E, H Schnell, P Rosenkranz (2011). Invasion of Varroa destructor mites into mite-free honey bee colonies under the controlled conditions of a military training area. Journal of Apicultural Research, 50(2), 138-144.
[10] https://scientificbeekeeping.com/the-varroa-problem-part-16b/
[11] Greatti, M (1992) Op. cit.
[12] Mortensen, A, C Jack, J Ellis (2018). The discovery of Varroa destructor on drone honey bees, Apis mellifera, at drone congregation areas. Parasitology Research 117(10): 3337-3339.
[13] Pfeiffer, K & K Crailsheim (1998). Drifting of honeybees. Insectes Sociaux 45(2): 151-167.
[14] Bordier, C, et al (2017) Should I stay or should I go: honeybee drifting behaviour as a function of parasitism. Apidologie 48(3): 286–297.
[15] Cappa, F, et al (2016) Bee guards detect foreign foragers with cuticular chemical profiles altered by phoretic varroa mites. Journal of Apicultural Research 55(3): 268-277.
[16] Geffre, A, et al (2020). Honey bee virus causes context-dependent changes in host social behavior. Proceedings of the National Academy of Sciences, 117(19): 10406-10413.
[17] Poulin, R (2010) Parasite manipulation of host behavior. Advances in the Study of Behavior 41: 151-186.
[18] DeGrandi-Hoffman, G, F Ahumada, H Graham (2017). Are dispersal mechanisms changing the host–parasite relationship and increasing the virulence of Varroa destructor (Mesostigmata: Varroidae) in managed honey bee (Hymenoptera: Apidae) colonies? Environmental Entomology 46(4): 737-746.
[19] Sakofski, F (1991) Quantitative investigations on transfer of Varroa jacobsoni Oud. In Recent research on bee pathology. International symposium of the International Federation of Beekeepers Associations, Gent (Belgium), 5-7 Sep 1990.
[20] Neumann, P, et al (2000) Colony evaluation is not affected by drifting and worker honeybees (Apis mellifera L.) at a performance testing apiary. Apidologie 31: 67–79.
[21] R M Goodwin, et al (2006) Drift of Varroa destructor-infested worker honey bees to neighbouring colonies. Journal of Apicultural Research 45(3): 155-156.
[22] Cervo, R, et al (2014) High Varroa mite abundance influences chemical profiles of worker bees and mite-host preferences. J. Exp. Biol. 217: 2998– 3001.
[23] Bordier, C, (2017) Op. cit.
[24] Peck DT & TD Seeley (2019) Mite bombs or robber lures? The roles of drifting and robbing in Varroa destructortransmission from collapsing honey bee colonies to their neighbors. PLoS ONE 14(6): e0218392.
[25] Peck DT (2019) op. cit.
[26] Kulhanek, K, et al (2021) Accelerated Varroa destructor population growth in honey bee (Apis mellifera) colonies is associated with visitation from non-natal bees. Scientific reports, 11(1): 1-15.
[27] Frey, E (2011) Op. cit.
[28] Kulhanek, K (2021) op cit.


