
The Varroa Problem: Part 16b – Bee Drift and Mite Dispersal (cont.)

October 28, 2018
Contents
Bee Drift and Mite Dispersal (continued) 1
So why do colonies allow bees to drift in?. 1
The amount of mite drift into other hives. 5
What happens to all the mite-infested bees when a colony collapses?. 8
Swarms coming back to bite you in the Butt. 9
The Varroa Problem: Part 16b
Bee Drift and Mite Dispersal (continued)
First Published in ABJ April 2018
Randy Oliver
ScientificBeekeeping.com
It’s clear that there may be a considerable amount of hive-to-hive drifting by bees and mites in the typical apiary. This then raises the question of whether there is anything that we can do about it?
So why do colonies allow bees to drift in?
Let’s start with some definitions. As I’m sure that you’re aware, the female members of the honey bee colony allocate their work to two distinct morphological castes–the fertile queen and the reproductively-suppressed workers. The workers further divide up their labor between behaviorally and/or physiologically distinct subcastes [[1]]. Some years ago, three heavyweight researchers in bee behavior–Breed, Robinson, and Page [[2]]–demonstrated that there are distinct guarding and soldier behavioral castes [[3]]. The guards are typically younger than the soldiers and station themselves at the nest entrance(s). The soldiers don’t fly much, but instead appear to “hang out” on the combs, perhaps waiting for a signal of alarm pheromone from the guards to stir them into action [[4]] to drive off a large intruder (by using warning bumps and/or and stinging).
Practical application: I’m explaining this so that the reader understands the difference between guarding behavior (which takes place at the hive entrance and periphery of the cluster), and soldiering behavior (the sort of mass stinging response to disturbance that beekeepers generally find distasteful). This may be an important distinction, since we may be able to breed for bees with good guarding behavior, without breeding for increased soldiering behavior.
Guard bees are responsible for warding off potential robbers attempting to enter the hive to steal the colony’s precious food stores, or other insects (such as hive beetles or yellowjackets) interested in the protein-rich beebread or brood. Guarding is also important to prevent “social parasitism,” such as that practiced by the Cape bee, in which a foreign “queen” can enter a hive and take over reproduction (similarly, a few bees in a “usurpation swarm” can enter a hive and overthrow the queen [[5]]). Just as a watchful police force is of necessity in human societies that hold items of value, the guard caste is of paramount importance for protecting the resource-rich honey bee nest from thieves and predators (Fig. 6).

Figure 6. The guarding subcaste is the first line of defense at any opening to the resource-rich nest cavity. These guard bees are harassing a foreign worker that drifted, and may not allow her to enter their hive. Should the guards emit alarm pheromone, that cue would then trigger the soldier bees inside to launch a defensive attack.
Worker bees typically engage in guarding behavior when they are around two to three weeks of age [[6]]. There is a genetic component to the proportion of the worker population that act as guards, as well as in determining their persistence [[7]]. Guard bees normally reject non-nestmates attempting to gain entrance to the hive—recognizing them by smell or by their behavior [[8], [9], [10]].
The importance of foreign worker rejection due to their exhibiting the wrong odor is even more important since the invasion of varroa –in an intriguing recent study out of Italy, the researchers discovered that the odor (volatile cuticular hydrocarbons) of adult workers changes when they are parasitized by a phoretic mite. Guard bees mounted a greater defense against previously parasitized workers than against those that hadn’t carried a mite [[11]].
Practical application: perhaps we could breed for this trait, thus helping to prevent invasion of mites into a hive.
Keep in mind, however, that allocating a proportion of the worker caste as guards comes at a cost to the colony as a whole. As explained in a nice overview by Rivera-Marchand [[12]]:
The cost of defense is minor for Africanized bees in the mainland where resources are abundant. These bees can afford to defend instead of storing food, because flowers are available through the year. However, for temperate bees… defense can affect [colony] growth… where resources are limited, it seems that defensive colonies are not able to gather enough resources for reproduction. Colonies with low defensive levels may have a reproductive advantage over the more aggressive colonies …”
Practical application: we can only hope that the fitness advantage gained by the extreme defensiveness of the Africanized bees may be lost as African alleles introgress northward into the established European populations in North America. This may, however, be wishful thinking, since the Western European (“German Black”) feral bees already established in some areas have long been known for their “nastiness” [[13]]. Only time will tell…
In order to overcome the costs involved in defense, European races of honey bees adjust the number of engaged guards conditional on the state of attractivity of their stored resources to competitors—that is, they set plenty of guards when they are protecting their stores from robbers, but once the nectar flow begins in earnest (and robbing pressure presumably decreases), the number of guards at the entrance decreases, and the rejection rate of foreign bees drops to near zero [[14]].
Butler and Free [[15]] found that the appearance of bees exhibiting the characteristic swaying-bobbing flight of robbers (which to a human appears “sneaky”) alerts the guards and increases the amount of guard bee interception of foreign bees. Ditto for the introduction of foreign intruders into the hive. Perhaps surprisingly, they also observed marked bees switching back and forth between guarding, foraging, and robbing. And as anyone who has spent time watching the hive entrance has seen, drifted foragers that act either “self confident” or “submissive” are generally allowed free passage by the guards; if instead they attempt to escape, the guards will grab them and try to sting them. If a drifted foreign bee spends 2-3 hours in a hive, it picks up enough of its new colony’s scent that it is no longer challenged by the guards of that hive.
Since drifting workers may be foragers intent upon robbing, or may carry infectious parasites or pathogens [[16]], it’s surprising to me that guards ever allow any drifting bees into a hive. However, it’s easy to observe that unless there is a nectar dearth, colonies tend not to exhibit many guards [[17]], and drifted bees are typically readily accepted. I suspect that this may have more to do with our unnaturally dense aggregation of hives in apiaries, rather than an innate willingness of colonies to accept foreign workers into their nest.
Investigating this further, I found a couple of studies of interest. One [[18]] found that ant colonies increased production of their soldier caste when the foragers were exposed to foreign ants. Similarly, Butler [[19]] found that introducing foreign bees at the entrance increased the amount of guarding, “suggesting that colonies regulate guard numbers in response to robbing and intrusion.”
On the other hand, Rittschof [[20]] found that colonies repeatedly experimentally disturbed exhibited less guarding behavior—apparently becoming habituated to the disturbance. Perhaps this also occurs in response to the continual drift occurring in an apiary.
The last question in my mind is related to the near-universal desire for beekeepers to breed for gentleness:
Practical breeding question: as much as I love to be able to work my sweet-tempered bees in shorts and tee shirt, has my selection for “gentleness” reduced their propensity to guard the entrance against foreign bees?
So much for between-hive drifting of bees, which certainly accounts for some degree of mite diffusion in an apiary. But far more mites appear to disperse during late-season collapse and robbing.
The sheer numbers involved
Sadly, for the vast majority of colonies in which varroa is inadequately managed, the combination of stress due to the mites, coupled with an in-hive epidemic of one or more viruses, ultimately results in that colony’s unfortunate collapse. At that time, there can be a lot of mites in the hive (Fig. 1).

Figure 1. The above simulation reflects the kind of alcohol wash counts [[21]] that I commonly see in untreated hives stocked with non-resistant bees; to the right are the numbers of mites necessary to produce such counts. That 17,000 figure may sound high, but is supported by simple math and hard data.
Colonies with very high mite numbers often collapse during the late summer or autumn, leaving behind a hive devoid of bees, but full of honey (Fig. 2).

Figure 2. Typical signs of a colony that has collapsed due to a varroa-vectored epidemic of DWV. Note the white fecal deposits from the mites, the bee with deformed wings, and the workers unable to emerge from their cells. The hive may be full of honey, but you’ll rarely see a mite.
So here’s the question: I don’t see any mites remaining in a hive after it’s collapsed—where the heck did they go? Mites don’t just walk out of a hive–they hitch a ride on an exiting bee.
An unanswered question: there are three main possibilities as to where each mite ends up:
- The bee carrying it flies off toward the horizon, and they both die.
- The bee carrying it drifts to another hive.
- The mite hops onto a robber bee to be carried to its
Obviously, options 2 and 3 have significant ramifications for other hives in the area.
The amount of mite drift into other hives
There have been a few studies in which researchers have quantified the amount of mite immigration into monitored hives in an apiary. I put their published data into the chart below (Fig. 3).
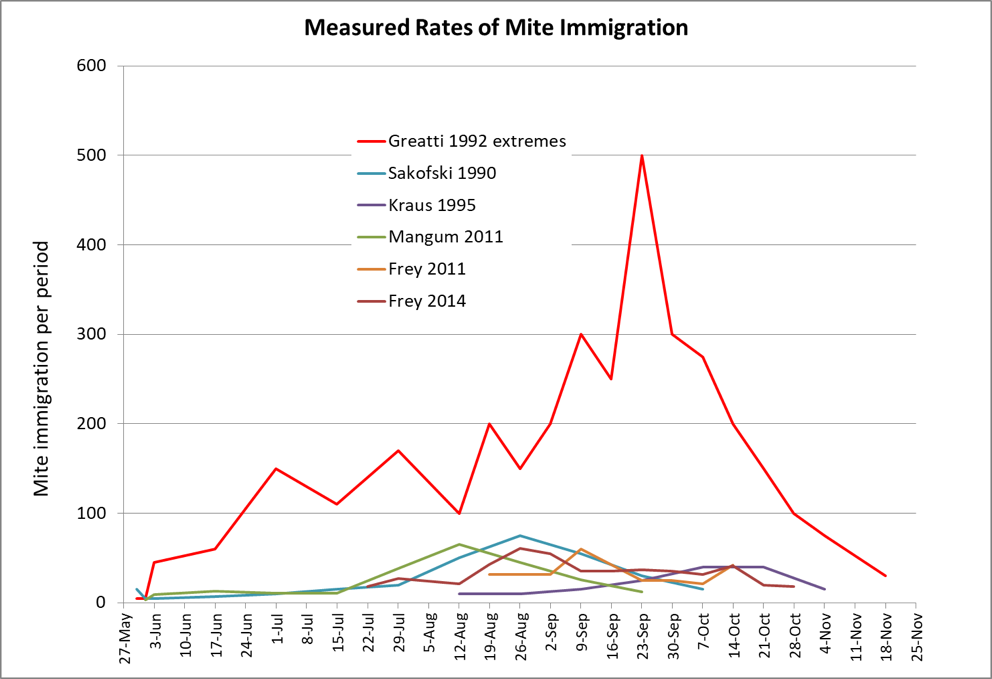
Figure 3. It is apparently not unusual for a colony to experience serious late-season varroa immigration numbering in the hundreds, and sometimes in the thousands of mites, as in the Greatti’s study, for which I used the most extreme values from their monitored hives. Data from [[22]].
The hard data above indicate that mite immigration is typically pretty low early in the season, and then spikes when there is late-season robbing and collapse. Although the counts by Greatti were substantially higher than those of other observers, they may more closely reflect what many of our colonies are experiencing these days.
Greatti counted between 2,000 and 3,500 mites invading each of their 10 monitored hives over the course of a season—with nearly 500 mites invading one hive over the course of 4 days, despite the fact that all surrounding apiaries either had been, or were undergoing, treatment with miticides.
Practical question: is this immigration of mites due mainly to the drifting of workers abandoning their collapsing hive, or instead due to bees engaged in robbing inadvertently carrying them home? And if not the bees in managed apiaries, is it from collapsing feral colonies and/or escaped swarms?
I have yet to see data that answers the above questions. Some research is already underway, and I hope to run an experiment myself this summer in order to help to answer the question. I’ll review some intriguing observations below.
Collapse and Robbing
As a colony begins to collapse, its virus-weakened workers no longer mount a defense against robbing bees; if there is a nectar dearth at the time, other colonies will help themselves to that unguarded honey. Some proportion of the mites in the dying colony would be expected to climb onto those robbers prior to their returning home.
What I found of great interest were the following observations by the Greatti:
It was difficult to discover the source of the mites: the question arises, where mites survive during acaricide treatments, since all the apiaries in the neighbourhood had been treated the year before, and again during September and October, so infestation levels of these colonies were low to moderate. Thus, it seems likely that most mites came from feral swarms. Only two feral colonies were detected near the experimental apiary (about 200 m from it); one was caught, the second did not survive repeated robbing.
After I published Donald Aiken’s data on his mite drops while he was applying oxalic acid vaporization (totaling some 17,000 mites from a single hive), he shared an observation:
The hive I treated and recorded the resulting mite drops from was very strong and produced over 200 lbs of honey. It actually continued to bring in honey after all the flowers were gone from my immediate neighbourhood. I conjectured at the time that they were sucking it out of the fence posts, but in retrospect they must have been robbing out weak hives that were infested with varroa.
Think about the above. Let’s say that foragers robbing a collapsing hive carry fairly heavy loads of honey (50 mg) back home. That would mean that every pound of weight gained by that hive would require 9,000 bee return trips from the hive being robbed. If those returning foragers brought back only 1 mite per every 10 trips, that would still be 900 new mites being brought home for every pound of honey gained!
I suspected that massive mite immigration occurred in some of my monitored hives last fall [[23]], but upon further modeling, I’m now not so sure—I need to monitor actual immigration to confirm.
Research needed: Donald’s observation gives me an idea for quantifying the correlation between robbing and immigration of mites into the robbing hives. In an area such as mine, where there is no natural colony weight gain late in the season, set up a bunch of treated hives on scales, and monitor the mite influx. Then look for a correlation between late-season weight gain and the increase in varroa immigration. This might help to clarify why only some colonies exhibit late-season spikes in mites. If you’re interested in helping to collect such data, see [[24]].
Greatti had another interesting observation [[25]]:
[In only] a few instances was robbing of infested colonies in hives or feral colonies observed… perhaps robbing does not always lead to massive attacks, particularly when other sources of forage are present, and thus it is difficult to observe.
Original from Greatti: “only in a few instances, robbing of infested colonies in hives or feral colonies was observed…Perhaps robbing does not always lead to massive attacks, particularly when other sources of forage are present, and thus it is difficult to observe.”
Greatti had another interesting observation—that in only a few instances was robbing of infested colonies in hives or feral colonies observed, and that perhaps robbing does not always lead to massive attacks, particularly when other sources of forage are present, and thus may be difficult to observe.
The above observation brings to mind Wyatt Mangum’s fascinating article on surreptitious robbing [[26]]. Could it be that as colonies begin to weaken from the varroa/virus complex, this sort of surreptitious robbing allows mites to rapidly disperse to new hives? Remember the previously-mentioned finding by Cervo that mites in highly-infested colonies may be attracted to the odor of foreign bees.
Practical application: prior to varroa, it was evolutionarily adaptive for a colony to steal honey from a weakened neighbor. But today that serving of free honey will likely come with a side order of mites.
I’ve yet to see good data as to the distance that foragers will go to rob; Seeley [[27]] suggests that at the low density in Arnot forest, scant robbing occurs. On the other hand, he and Loftus [[28]] discuss observing a spike in mite infestation levels in hives when a colony 60 meters distant collapsed. But even once we figure out the range for robbing, I’m still left struggling with yet another question:
What happens to all the mite-infested bees when a colony collapses?
We’ve all seen it—an entire colony appears to “disappear” overnight. Since there are no dead bees to be seen, it’s clear that the entire workforce abandoned the hive. The as-yet-unanswered question then is, where do those bees (and all the mites that they’re carrying) go? Do they simply fly off towards the horizon to die? Or do some of them drift into other hives? There is surprisingly little data on this, although Dr. Dennis vanEngelsdorp has mentioned that one of his team found marked bees drifting from a collapsing colony into other hives at considerable distance.
Research needed: this is a crucial question crying for an answer—do all those mite-infested bees just “disappear,” or do they instead flood into surrounding hives? We clearly need far more data on the drifting of marked bees from collapsing colonies to other hives in order to understand why we observe such high rates of late-season mite immigration. It would be fairly simple to quantify the amount of drifting that occurs at collapse by stocking a yard with hives of yellow and black bee stocks, and allowing the hives of one color to collapse, then inspecting the hives of the other color to quantify the number of drifted bees during those collapse events. Please contact me if you’d like to run this important, yet simple experiment in one of your yards.
OK, it’s pretty clear that drift and/or robbing could account for large amounts of mite dispersal into other hives. But as pointed out by Greatti:
The degree of reinfestation of single colonies, on different days of observation, varied greatly; however, the most reinfested or the least reinfested colonies of a group often remained the same in successive observations, and sometimes for long periods of time…Marked differences among colonies of the same group in the same day of observation were often observed…
That’s exactly what I observed with the potential breeder queens that I was tracking last season. I observed huge mite spikes in only half my monitored hives in any apiary—the rest maintained low varroa levels, which then raises the question:
Selective breeding questions: some colonies appear to pick up a huge load of mites in early autumn, yet others somehow manage to maintain very low mite levels. Can we breed for colonies that prevent such mite immigration?
We may not yet know the answers to the above questions, but there is an aspect of apiary management that may be very helpful in minimizing autumn mite immigration.
Swarms coming back to bite you in the Butt
I often hear beekeepers shrug off, “yeah, I think that half my colonies must’ve swarmed this spring” (Fig. 4). But it’s not like those swarms just disappear once they fly off.

Figure 4. It’s easy to rationalize that it was just too hard to prevent your colonies from swarming. But any escaped swarm has the potential to become a “mite factory” whose output you may later come to regret.
Allow me to share one final quote from Greatti:
A lost swarm can be a source of thousands of mites at the end of the season or during the following year, and thus can endanger several colonies, when it may be weakened by heavy Varroa infestation and is consequently liable to robbing by other colonies.
We frequently hear of the propensity of Africanized bees to swarm frequently, but as pointed out by Medina-Flores [[29]], this strategy may be most adaptive in areas with short winters, regular nectar flows, and plenty of nesting cavities:
In the temperate dry region that we studied, blossoms are scarcer, colder temperatures reduce colony reproduction during winter, and its vegetation provides fewer nesting sites than in subtropical regions. Thus, it is less likely for bee colonies to swarm frequently and to establish feral populations in temperate climates than in more tropical environments.
Nevertheless, both Winston and Loftus [[30]] have documented that temperate colonies may swarm several times a season if they crowd their cavity. And the cold-adapted Primorsky Russian bees swarm at the drop of a hat. When conditions are good, it would be normal for most of the hives in an inadequately-managed apiary to swarm at least once per season.
Those swarms carry a starting population of mites with them, and if they are successful at establishing a nest, that varroa population will build up. Depending upon timing, that swarm colony will eventually collapse from the varroa/virus complex—typically with a population of approximately 5,000-15,000 hungry mites. The question is, how many of those mites will make it back into your hives when that collapse inevitably takes place?
Practical application: it appears that one of the most important mite management tools would be to minimize the number of swarms that issue from your own apiary. Remember that every lost swarm has the potential to create a nuisance with your neighbors, to become competition for your bees, and then a mite factory that will eventually collapse. Controlling swarming is good management!
Acknowledgements
Thanks as always to Pete Borst for research assistance, and to all the dedicated and hard-working bee researchers from whose publications I draw useful information.
Notes and Citations
[1] There are genetic components as to how each patriline of sisters behaves, as well as environmental cues and triggers.
[2] Breed, MD, GE Robinson, RE Page (1990) Division of labor during honey bee colony defense. Behav. Ecol. Sociobiol. 27: 395-401. Worth reading!
[3] Reviewed in: Nouvian, M, et al (2016) The defensive response of the honeybee Apis mellifera. Journal of Experimental Biology 219: 3505-3517 doi:10.1242/jeb.143016 Open access.
[4] Suggested by Breed, and supported by my own observations.
[5] https://scientificbeekeeping.com/whats-happening-to-the-bees-part-2/
[6] Winston, ML (1987) The Biology of the Honey Bee. Harvard University Press.
[7] Breed MD, Rogers KB (1991) The behavioral-genetics of colony defense in honeybees—genetic-variability for guarding behavior.. Behav Genet 21:295–303.
[8] Dani, F, et al S (2005) Nestmate recognition cues in the honey bee: differential importance of cuticular alkanes and alkenes.. Chem Senses 30:477–489.
Of interest is that attempting to mask the “foreign” smell with an added odorant may not be effective.
Couvillon, MJ, FLW Ratnieks (2008) Odour transfer in stingless bee marmelada (Frieseomelitta varia) demonstrates that entrance guards use an “undesirable–absent” recognition system. Behavioral ecology and sociobiology 62, Issue 7, pp 1099–1105.
Ratnieks FLW, et al. (2011) Acceptance by honey bee guards of non-nestmates is not increased by treatment with nestmate odours. Ethology 117: 1–9.
[9] Breed, MD, et al (1998) Comb wax effects on the ontogeny of honey bee nestmate recognition. Anim Behav 55:13–20.
[10] Well reviewed by Breed, MD, et al (2015) Chapter 9 Nestmate Recognition in Eusocial Insects: The Honeybee as a Model System in L. Aquiloni and E. Tricarico (eds.), Social Recognition in Invertebrates.
[11] Cappa, F, et al (2016) Bee guards detect foreign foragers with cuticular chemical profiles altered by phoretic varroa mites, Journal of Apicultural Research 55(3): 268-277.
[12] Rivera-Marchand, B, et al (2008) The cost of defense in social insects: insights from the honey bee. Entomologia Experimentalis et Applicata 129: 1–10.
[13] The dark Apis mellifera mellifera bloodline (M mitotype) established itself as feral populations the U.S. early on, This is one reason that the “gentler” Italian and Carniolan (C mitotype) bloodlines were more popular. Many of us older beekeepers remember the “hot” dark feral bees that used to be common, and the studies by Magnus and Szalanski (https://scientificbeekeeping.com/whats-happening-to-the-bees-part-5-is-there-a-difference-between-domesticated-and-feral-bees/) confirm that the M line of bees still thrives in some areas.
[14] Downs SG, Ratnieks FLW (2000) Adaptive shifts in honey bee (Apis mellifera L.) guarding behavior support predictions of the acceptance threshold model.. Behav Ecol 11:326–333.
[15] Butler CG, Free JB (1952) The behaviour of worker honeybees at the hive entrance. Behaviour 4:262–292.
[16] As noted by Free, the “infrequency, or absence, of drifting of bees between wild colonies may be one reason why they are comparatively free from disease.”
[17] At least with the gentle bee stock the I keep; I’m curious as to the degree of guarding exhibited by wild-type bees. Please let me know if you have observations.
[18] Passera, L, et al(1996) Increased soldier production in ant colonies exposed to intraspecific competition. Nature, 379, 630-631.
[19] Butler CB, Free JB (1952) The behaviour of worker honeybees at the hive entrance.. Behaviour 4:262–292.
[20] Rittschof, CC & GE Robinson (2013) Manipulation of colony environment modulates honey bee aggression and brain gene expression. Genes, Brain, and Behavior 12(8): 802–811.
[21] Per half cup of bees.
[22] I either took published numbers from tables, or scaled data off of published graphs. Since the various authors collected data over different time intervals, I needed to take some liberties in adjustment in order to create the graph.
Sakofski, F, et al (1990) Seasonality of honeybee colony invasion by Varroa jacobsoni Oud. Apidologie 21:547–550
Greatti M, Milani N, Nazzi F (1992) Reinfestation of an acaricide-treated apiary by Varroa jacobsoni Oud. Exp Appl Acarol 16:279–286
B Kraus, Re Page Jr. Population growth of Varroa jacobsoni Oud in Mediterranean climates of California. Apidologie, Springer Verlag, 1995, 26 (2), pp.149-157.
Mangum, WA (2011) Varroa immigration and resistant mites. ABJ 151(5): 475-477.
Frey E, Schnell H, Rosenkranz P (2011) Invasion of Varroa destructor into mite-free honeybee colonies under the controlled conditions of a military training area. J Apic Res 50:138–144.
Frey E, Rosenkrantz P (2014) Autumn invasion rates of Varroa destructor (Mesostigmata: Varroidae) into honey bee (Hymenoptera: Apidae) colonies and the resulting increase in mite populations. J Econ Entomol 107:508–515.
[23] See my article “Selective Breeding for Mite Resistance,” which should be posted to my website by the time of publication of this piece.
[24] https://scientificbeekeeping.com/suggested-protocol-to-determine-amount-of-mite-immigration/
[25] I slightly paraphrased this quote to make it read better.
[26] Mangum, W (2012) Robbing: Part 2: Progressive robbing. ABJ 152(8): 761-764.
[27] Seeley, TD (2007) Honey bees of the Arnot Forest: a population of feral colonies persisting with Varroa destructor in the northeastern United States. Apidologie 38: 19–29.
[28] Loftus JC, ML Smith, TD Seeley (2016) How honey bee colonies survive in the wild: testing the importance of small nests and frequent swarming. PLoS ONE 11(3): e0150362.
[29] Medina-Flores CA, et al 2014. Africanized honey bees (Apis mellifera) have low infestation levels of the mite Varroa destructor in different ecological regions in Mexico. Genet.and Mol. Res. 13:7282-7293.
[30] Winston, ML (1987) The Biology of the Honey Bee. Harvard University Press.
Loftus JC (2016) op cit. (and Seeley, pers. comm.)


