
Understanding Colony Buildup and Decline: Part 9b – The Regulation of Bee Longevity

January 29, 2017
Understanding Colony Buildup and Decline: Part 9b
The Regulation of Bee Longevity
Randy Oliver
Scientific Beekeeping
First Published in ABJ in February 2016
In the first part of this article, I looked at the regulation of the lifespan of the long-lived “winter” (diutinus) bees. Now it’s time to turn to the question of why “summer bees” live such short lives.
Update 25 July 2018:
An interesting paper was recently published–the title describes its contents:
Effects of flight activity and age on oxidative damage in the honey bee, Apis mellifera. J Exp Biology doi: 10.1242/jeb.183228
The authors found evidence that the intense muscular activity of flight leads to more rapid senescence, but also discusses that more study is needed on diutinus bees. The authors don’t mention it directly, but the question to me is how diutinus “winter” bees, some of which engage in extended use of their flight muscles to produce heat in the insulating shell of the winter cluster, apparently manage to live for hundreds of days.
Another study of interest is: Rueppell, O, et al (2007) Aging without functional senescence in honey bee workers. Curr Biol. 17(8): R274–R275.
Another study, on the other hand in support of ingrained natural senescence, concluded that “We have shown here that limited worker lifespan is due at least in part to intrinsic senescence and not solely to extrinsic mortality factors.” They concluded that a worker’s warranty was for roughly 50 days, independent of use.
Remolina, S, et al (2007) Senescence in the Worker Honey Bee Apis mellifera. J Insect Physiol. 53(10): 1027–1033.
The Last Piece of the Puzzle
There yet remains one answered question. Keeping in mind Ruepell’s observation that bee aging patterns appear to be under strict regulatory control, I can’t help but wonder what determines “normal” worker longevity. Why, during the spring and summer, do worker bees exhibit such an abbreviated, yet consistent, average longevity of only 35 days? It hardly seems that evolution would leave summer worker longevity to chance.
The most common explanation is that those summer bees simply wear themselves out by foraging. But does that explanation hold up to closer scrutiny?
Foraging is indeed high risk behavior, what with wind, rain, predators, and problems with orientation. Up to a third of workers do not successfully return from their first foraging flight [1]. A number of researchers have found that once a worker starts foraging, their remaining life expectancy is generally only 8-12 days [2]. Workers tend to live shorter lives during intense honey flows, likely due to wing wear and the oxidative stress associated with the hard work. And it’s clear that the earlier a worker initiates foraging behavior, the shorter its expected overall lifespan. So is foraging then the cause of the short lifespan of summer bees?
There are a couple of pieces of evidence that make me question this hypothesis [3]. Firstly, those long-lived winter bees also use their flight muscles to generate heat, yet still live a long time. And secondly, a well-designed study by the legendary experimental ecologist Paul Schmid-Hemple [4] found that restricting the amount of flight taken by foragers did not increase their average lifespan beyond 40 days (although the harder they worked, the shorter their lifespan). A later study [5] supported this finding (with mortality of either foragers or non-foraging workers generally increasing sharply at around 45-50 days of adult life.
Working conclusion: these two studies appear to strongly refute the hypothesis that forager longevity is determined solely by wear and tear. Perhaps we should look more closely at the reason that summer bees live such short lives.
Update: A relevant quote from:
Becerra-Guzmán, F., Guzmán-Novoa, E., Correa-Benítez, A., & Zozaya-Rubio, A. (2005). Length of life, age at first foraging and foraging life of Africanized and European honey bee (Apis mellifera) workers, during conditions of resource abundance. Journal of apicultural research, 44(4), 151-156.
“What is not clear is if senescence depends on foraging activity. Whether senescence depends onactivity or not is crucial for understanding the evolution of division of labour in social insects.”
Let’s think about what sets the lower and upper limits on the longevity of a summer bee. The lower limit is set by three constraints:
- The minimum amount of time that it takes a newly-emerged bee to develop her brood food glands (~3 days),
- Spending enough time in nursing duties to rear not only a replacement bee, but also perhaps two additional workers to allow for colony growth [6], as well as providing jelly to the queen and foragers (let’s estimate a 7-10 day minimum).
- A forager’s expected longevity once it initiates foraging (8-12 days).
From the above, I come up with a minimum required longevity of around 20-23 days (although the lifespan can be even shorter if a worker curtails nursing duties and initiates precocious foraging). So how about the upper limit? Haydak [7] found that when nurses were forced to continue feeding brood (by experimentally removing sealed brood prior to emergence), that they could still produce jelly for at least 75 days, and live for at least 135 days. It appears that their maintenance of a high level of vitellogenin in their bodies minimizes natural senescence. And it also appears that the intense production of jelly for a prolonged period does not necessarily cause premature aging.
My question: remember the evolutionarily-determined strict regulatory control of aging? Now that we’ve set the lower (20-23 days) and upper (>135 days) limits on summer bee theoretical or experimentally observed longevity, the question then is: Why has evolution narrowed the normal lifespan of summer bees to be only 35 days (rarely reaching 60 days) despite their potential for living much longer?
The colony gains great benefit from the nurses. They function as the digestive system for the colony, and act as surrogate mothers for every bee (queen, larvae, foragers, drones) needing nourishment. The colony protects them, keeping them safe in its confines.
After completing her requisite jelly production duties, the now mid-aged nurse gets pushed off the brood combs by younger nurses, and shifts to being a “general purpose” mid-aged (“house”) bee. As she tapers off from processing pollen, she’ll move to processing nectar, building comb, ventilation, guard duty, etc. But at this mid-aged stage, due to her relatively high level of vitellogenin, one would expect her to age rather slowly (as observed by Haydak). So why don’t all summer bees simply spend additional time in the hive, and thus prolong their lives—thereby allowing the colony to keep increasing its population towards the goals of either reproduction or exploiting the main nectar flow?
Two reasons come to mind:
- The colony doesn’t require any more nurses or mid-aged bees, since those duties are filled by the subsequent emergence of new workers. If the idle mid-aged bees prolonged their lives in the hive, at some point they’d become a burden to the colony. So, having fulfilled their in-hive duties, summer bees then graduate to risky foraging.
- It may actually be to the colony’s detriment for summer bees to live for a longer period of time than 35 days on average. This may seem counterintuitive, but give me a moment.
I’ve spent countless hours mulling over this question. And my tentative conclusion is that the 35-day mean lifespan of foragers was determined by striking a balance between a longevity that is long enough to allow a colony to grow at an extraordinary clip during the growth and reproductive phases, and to store adequate reserves during the food storage phase, yet short enough to quickly dump any excess worker population when called for during conservation mode (in times of pollen dearth). To test this hypothesis, allow me to model the rates of colony population growth or decline when I change the variable of the worker survivorship curve.
The Effect of Worker Longevity on Colony Buildup
In order to see whether extending worker longevity would be of benefit during the colony growth, reproductive, and food storage phases, I plotted out simulations of colony growth with queens laying a realistic 1000 eggs/day, starting with broodnests already filled to the queen’s laying capacity (Fig. 1):
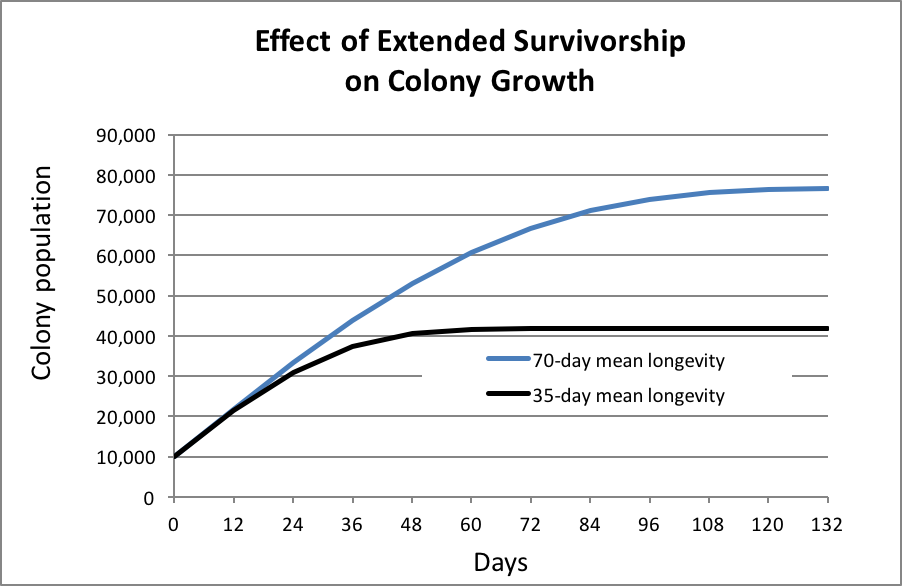 In the simulation above, increasing worker survivorship did indeed allow the colony to grow larger, but would that be of adaptive benefit? Keep in mind that colonies typically swarm when they reach a population of around 20,000 (Winston [9]) to 40,000 bees (strong colonies returning from almonds, pers. obs.). This simulation suggests that there would be only small benefit of extended survivorship of the workers during early colony growth and reproduction. Nevertheless, the colony has a couple of tricks up its sleeve to take advantage of that benefit.
In the simulation above, increasing worker survivorship did indeed allow the colony to grow larger, but would that be of adaptive benefit? Keep in mind that colonies typically swarm when they reach a population of around 20,000 (Winston [9]) to 40,000 bees (strong colonies returning from almonds, pers. obs.). This simulation suggests that there would be only small benefit of extended survivorship of the workers during early colony growth and reproduction. Nevertheless, the colony has a couple of tricks up its sleeve to take advantage of that benefit.
A question: It would likely be adaptive for workers to live a bit longer during the growth and reproductive phases of colony development. Remember, it’s pitch dark and crowded in the broodnest. So how could a worker assess how large her colony had grown?
This question may have been answered by Pankiw [10], who found that higher titers of queen mandibular pheromone (QMP) caused a 3-day delay in the age at which workers transitioned to foraging. The titer of a queen’s output of QMP in the workers is proportionate to the number of bees in the hive [11]. This pheromonal regulatory signal would have the effect of extending worker contribution to in-hive tasks, as well as their overall longevity. Both effects would likely be of benefit to growing colonies. And those effects would diminish as the colony became more populous, thus being self regulating.
And as far as ability to store honey, the potentially larger maximum population would not occur until relatively late in the season, and thus be of little benefit in areas in which the main nectar flow ends before that time. My conclusion: I don’t see a strong benefit to the typical colony of extending worker longevity during the growth, reproductive, or food storage phases [12]. So this leaves me to look at its effect during the conservation mode.
Selective Pressure During Conservation Mode
Once a colony has swarmed and then refilled its combs with honey, there would be no reason for it to maintain an idle and hungry workforce larger than that necessary to protect the queen and honey reserves. That takes only 5-10 thousand bees. The remaining thousands of workers in the hive would be superfluous—requiring more resources to maintain them than they’re worth.
We should first remember that bee behavior (including the regulation of senescence) evolved when bees lived in small tree cavities. A typical cavity (about the volume of a single Langstroth deep; larger for northern races, smaller for Italians) would hold only about 20,000 bees [13], and those colonies would normally swarm whenever the population reached that number. Thusly, bee behavior evolved long before beekeepers artificially increased the cavity size. Our domesticated stocks have been artificially selected against swarming behavior—my own year-old queens rarely swarm until the colony fills two deeps (around 35,000 bees). We should keep in mind that the basics aspects of bee biology evolved to be adaptive for colonies that seldom exceeded a population that would fill a single deep. I’m not sure how much we’ve changed bee behavior to account for our much larger hives and colony sizes.
Practical application: Also consider that our artificially-selected domestic stocks (bred for commercial beekeeping) are in general not as environmentally responsive as are more “wild type” temperate stocks, such as Russians or Carniolans, whose colony populations swing up or down rapidly in response to pollen availability. This could be one of the reasons why our domesticated Italian-type stocks, which are adapted to receiving feeding and other husbandry, do not survive in the wild as well as do some feral stocks [14]. I’m not in the least criticizing our high-production domestic stocks— they are bred for a purpose. Just don’t expect them to survive without some degree of husbandry—what we call bee-keeping.
Back to the question of why workers live for only 35 days, there doesn’t appear to be much benefit (nor detriment) for greater longevity during the buildup, reproductive, or food storage phases of colony dynamics. This suggests that we should look for an adaptive benefit during the conservation mode. I suspected that it had something to do with the cost of maintaining an excessive workforce.
Key point: in typical natural landscapes in temperate climes, a colony is in the growth, reproductive, and food storage phases for only a few months of the year (Fig. 2). During that time it applies its resources to go full tilt boogie in rearing as many bees as it can to increase its population—the more workers the better. This strategy completely changes when the colony goes into conservation mode—now its focus is upon simple survival through the pollen dearth and winter cold. To that end, it must carefully conserve its hard won honey stores.
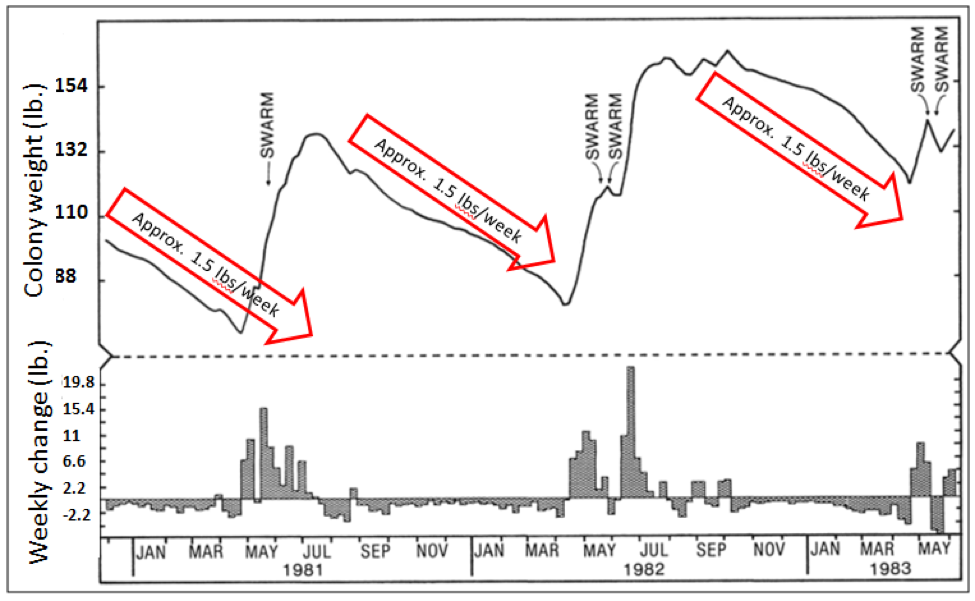 Part of the weight loss of the above colonies was from a reduction in worker population, but that would only account for 5-10 lbs overall. That leaves honey consumption—the more mouths to feed, the more honey used. The fact that the colonies lose weight from midsummer on indicates that the workforce is consuming more honey than it brings in.
Part of the weight loss of the above colonies was from a reduction in worker population, but that would only account for 5-10 lbs overall. That leaves honey consumption—the more mouths to feed, the more honey used. The fact that the colonies lose weight from midsummer on indicates that the workforce is consuming more honey than it brings in.
I suspected that the colony would want to dump the excess workers left over from colony peak population during the food storage phase (the main flow) as quickly as possible, in order to avoid having to feed them. But even if the nurse bees immediately ceased rearing brood, that wouldn’t have any effect on the adult worker population for three weeks. And we don’t see workers excluding other workers from reentering the hive (the fate of unwanted drones). But one way for the colony to quickly dump excess workers would be if they exhibited programmed early death.
Modeling the Effect of Longer Survivorship
Curious to see to what degree the survivorship curve of the workers effects the reduction of population after the main flow, I modeled what would happen if conditions suddenly turned bad, and a colony stepped hard on the brakes by completely ceasing broodrearing (Fig. 3):
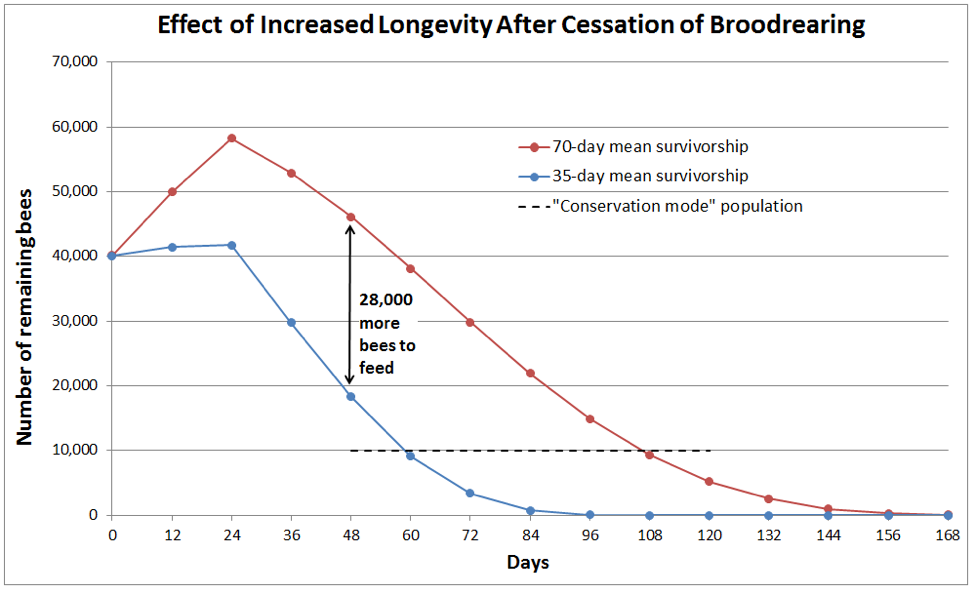 In answer to my question, the colony with extended survivorship would have thousands more bees to feed until the worker population eventually dwindled to the 10,000-bee level (a 5-6 frame cluster), and it would take 48 days longer to reach that population.
In answer to my question, the colony with extended survivorship would have thousands more bees to feed until the worker population eventually dwindled to the 10,000-bee level (a 5-6 frame cluster), and it would take 48 days longer to reach that population.
Similar to maintaining an army or idle workforce, there is a cost for keeping unnecessary workers in the hive. So I calculated the total amount of extra “bee days” between the two curves until they each reached the dashed line. And from that figure, based upon the resting metabolic rate of idle bees, I could arrive at the cost to the colony to maintain them [17]. It worked out to be an extra 58 lbs of precious honey consumed—honey that would not then be at hand for winter survival. Clearly, there is strong adaptive benefit for the honey bee to include programmed early death in its suite of colony population regulation mechanisms.
My proposed hypothesis: if foragers didn’t die at an early age, the workforce built up for the honey flow would become a liability afterward. Nature has perhaps selected for programmed early death of summer bees in order to allow the colony to quickly reduce its population, when necessary, in order to conserve honey stores. The typical 35-day mean survivorship has likely been worked out through thousands of generations of selective pressure as to be the best balance between the capability of the colony to grow rapidly, versus its means to quickly shed excess workers when they are unneeded.
Senescence vs. Death From Foraging
There’s a difference between death due to senescence, and death due to engaging in risky behavior. Bees appear to use both forms in order to regulate worker longevity. I’ve already mentioned two studies that indicate that there appears to be an ultimate limit to worker age during the summer, with them giving up the ghost at around 40-50 days of adult life, no matter whether they engage in foraging or not. Neukirch similarly found that in a colony allowed access only to a small flight cage, mean worker longevity was around 55 days, despite some living for 30 days after initiation of foraging in the cage. Thus, I suspect that there is some sort of programmed senescence that absolutely limits summer bees to a relatively short longevity, independent of whether they forage or not.
However, if a worker does start foraging, it typically dies in around 8 days. Although Neukirch and others have proposed that this was due to the exhaustion of some physiological aspect of the flight muscles, oxidative stress, or wing wear, Visscher and Dukas [18] found that foragers die at an exponential rate, suggesting that it was predation or other risks, rather than senescence, that limited their lifespans. However, their data was for tiny colonies in observation hives (~3000 bees per hive), which, as subsequent research has found, would be expected to have very different titers of queen and brood pheromones than full-sized colonies, so there could have been something else going on (I’m cautious about extrapolating all findings from studies of tiny observation hives to what goes on in full-sized colonies).
It’s plausible that the nurse bees can help to extend forager longevity when colonies are very small, or during times of dearth during spring buildup. In correspondence with Dr. Tom Seeley just before sending this article off to press, he speculated that:
What this means, I think, is that worker bees that have become foragers are not locked in to having short lives. In the early summer, at least, when a colony still needs to acquire lots of food, foragers can have long lives when they don’t have a lot to do, perhaps because at this time of year and under these conditions they are fed proteinaceous food to keep them going? The key point here is that I think we have evidence that the transition to foraging does not necessarily put a worker on a track to a short life. Generally, foragers are on a track to a short life (often because foraging is risky), but it seems that in early summer, at least, colonies can treat foragers in ways that will help them live long lives (not just 8-12 days on average), esp. if little forage is available.
The above is supported by Nuekirch’s data, which suggests that workers live longer early in the season (when pollen is plentiful) as opposed to later in the season.
Honestly, we still have much to learn about the regulation of worker longevity, but it appears to me that although the colony may have ways of prolonging the survivorship of its foragers (other than them just being lucky), that the main regulatory determinant of worker survivorship is the age at which they transition to foraging. And that age appears to be strongly determined by the exposure to the pheromone produced by young larvae. When pollen is plentiful, there’s lots of young brood, and emerging workers get a strong hit of their pheromone. This appears to put that worker on track for shifting to dangerous foraging at around three to four weeks of age, and if those foragers are not fed well by the nurses, they will soon die from either risk or senescence. And even if well supported, programmed senescence appears to kick in at around 40-50 days.
Update: A recent paper discusses various mechanisms by which bees could use epigenetic DNA methylation in order to affect senescence [19].
The Power of the Nurses
The nurse bees are absolutely key to regulating both colony growth and population reduction, since it is they who control the currency of jelly (protein). When fresh pollen is abundant, they flood the colony with jelly, and it goes full tilt into growth and/or reproduction (swarming). During a major nectar flow, they may be forced to abandon broodrearing, and shift to nectar processing, comb building, or even early foraging (thus resulting in a lack of emerging workers three weeks later, which then helps to reduce the workforce).
Especially interesting is how the nurses can help to rapidly dump excess worker population during times of pollen dearth. They can cannibalize eggs and larvae (thus recycling the larval protein back into jelly for redistribution); this would stop further recruitment of workers starting in a couple of weeks. Or they may restrict the amount of jelly that they feed to the remaining larvae, which would have the effect of shortening the lifespan of those workers when they finally emerge as adults [20]. They can also cut back on the amount of jelly that they share with newly-emerged workers, thus causing them to forage at an earlier age. And don’t forget that the existing foragers are dependent upon jelly begged from the nurses for their body maintenance; this mechanism could conceivably allow the nurses to accelerate the senescence of the foragers.
Pete Borst pointed out to me that I shouldn’t ignore the importance of a nectar flow. Although it is the availability of pollen that limits the nurses’ ability to produce jelly, it is a common observation that colony growth or decline (during warm weather at least) is also dependent upon at least a minimum of nectar income. I have not investigated how the presence or ingestion of nectar plays into aging.
Parsimony, Flexibility, and Redundancy
The Law of Parsimony suggests that the colony would regulate the physiological senescence of its members via a few simple triggers, and this indeed appears to be the case. The most important trigger appears to be the availability of fresh pollen—which leads to high levels of vitellogenin throughout the colony, due to the sharing of jelly produced by the nurses (using the colony’s “common stomach” as an information center [21]). This signals the colony members to go into growth mode, resulting in the release of another important signal—brood pheromone [22]. These simple signals alone allow the colony to rapidly respond to changes in the environment (mainly the presence, of lack, of incoming pollen). Of course, there are additional levels of regulatory redundancy, involving pheromonal and behavioral communications of information. Together, this complex sharing of information ideally strikes an optimal and ever changing balance.
The beauty of the system is its flexibility. Worker longevity can be greatly extended or shortened according to the colony’s needs. In the worst case—no incoming pollen, and thus no broodrearing–the newly-emerged forager will load itself with vitellogenin and become a long-lived diutinus bee, thus maximizing the queen’s chance of survival (keep in mind that every member of the colony other than the queen is expendable).
On the other hand, during good times, a worker goes through the “normal” progression of cleaner/nurse/house bee/forager. But this progression can be adjusted, as described eloquently by Maisonnasse [23]:
[The] queen, old brood and foragers produce pheromones…which slow down the progression of young bees towards the tasks typical of older bees. But young larvae also have something to say. [Their main pheromone has] the opposite effect of old brood on bee maturation, which is to accelerate worker age at first foraging. In this way, worker maturation occurs in a complex milieu of pheromonal compounds.
The pheromone produced by the young larvae not only accelerates the maturation of workers to forager status (thus shortening their overall longevity), but also tells the foragers that the broodnest is hungry for protein, thus causing them to preferentially forage for pollen [24]. And if pollen reserves run a bit short, that worker may engage in “precocious” foraging (greatly shortening its life). Conversely, if the colony is for some reason short on nurses, a nurse can prolong its nursing duties (and thus extend its life); and if the situation is desperate, mid-aged or early forager bees can (to some extent) even revert back to nursing duties.
Once pushed out of the broodnest by younger nurses, the ratio between mid-aged nectar processors and comb builders to foragers is regulated by the communication systems that I described in my previous article [25]. Mid-age bees (still enjoying delayed senescence due to their high level of Vg) can “hang out” for a while until needed for foraging, held back by the pheromone ethyl oleate produced by the foragers [26]. Again from Maisonnasse:
An overabundance of foragers leads to a lack of nurses in the colony, and thus a decline in brood care; conversely too many nurses cause a decrease in food storage in the colony and a subsequent decline in food for brood nourishment due to the scarcity of foragers. Consequently, a proper nurse-forager ratio is key to maintaining honey bee social homeostasis.
Upon taking up foraging behavior, the worker gives up any remaining Vg in its body to younger workers. By thus foregoing Vg-associated tissue repair, antioxidant, and immune functions, it gains greater efficiency as a forager, but at the cost of limiting its remaining lifespan—hence, programmed senescence. Yet even this can apparently be adjusted. Huang and Robinson [27] tracked juvenile hormone levels over the fall-to-spring transition (and when a colony was artificially chilled during summer), finding that titers of this “aging” hormone varied greatly with conditions. However, once a forager, longevity apparently can never be fully restored–even if prevented from foraging (say by inclement weather), the forager will still soon die of “old age.” As with stinging, or the altruistic suicide of sick bees [28], this sacrifice of potential longevity, although fatal to the individual worker, is of benefit to the colony (and thus the species) as a whole.
Closing Notes
The mechanisms of colony homeostasis are both elegant in their simplicity yet complicated in their necessary redundancy. The more that we understand bees, the better we can work with them.
Practical application: I cannot tell you how many times I’ve heard beekeepers explain why they did something for a reason that made sense to them, but in reality would have an entirely different effect from that expected, because the beekeeper didn’t understand bee behavior. Bees follow their own set of rules. By learning those rules, we can make better informed management decisions.
Nature has selected for an optimum longevity of the honey bee female phenotypes, which is longer for some and shorter for others. Perhaps natural selection will need to shift the survivability curves in response to the introduction of varroa, DWV, and Nosema ceranae.
Acknowledgements
I thank Lloyd Harris, Gro Amdam, and especially Tom Seeley for their constructive comments on this manuscript. I also wish to express my appreciation of the other pioneering researchers (as well as their many students) who have guided my way in my attempts to understand bee behavior and colony homeostasis: Knut Schmid-Nielson, Bernt Heinrich, Paul Schmid-Hemple, Mark Winston, Karl Crailsheim, Yves Le Conte, Gene Robinson, Zach Huang, Rob Page, and Olaf Rueppell (apologies to those I overlooked).
As always, I could not research these articles without the help of my collaborator Peter Borst. And of course thanks again to Lloyd Harris for his data set and helpful discussion, without which I would never have written this series.
Notes and Citations
[1] Thompson, H, et al (2015) Thiamethoxam: assessing flight activity of honeybees foraging on treated oilseed rape using RFID technology. Environ Toxicol Chem., Accepted Article • DOI: 10.1002/etc.3183
Rueppell, O, et al (2007) Regulation of life history determines lifespan of worker honey bees (Apis mellifera L.). Exp Gerontol 42(10): 1020–1032.
[2] Neukirch, A., 1982. Dependence of life span of the honeybee (Apis mellifera) upon flight performance and energy consumption. J. Comp. Physiol. 146 : 35 – 40.
Some workers during summer may be the odd leftover diutinus bee. Lloyd Harris recently wrote me that, “I marked/painted some more workers last September and to my surprise I had 1 worker bee that was still alive in mid-July. It lived at least 312 but less than 324 days. This is the longest lived worker bee that I have ever observed. It lived almost as long as those observed by Farrar.”
[3] Thomas Huxley’s “The great tragedy of science – the slaying of a beautiful hypothesis by an ugly fact.”
[4] Schmid-Hempel, P., Wolf, T., 1988. Foraging effort and lifespan of workers in a social insect. The Journal of Animal Ecology 57, 509–521.
[5] Rueppell, et al (2007) op cit.
[6] At maximum, a nurse can produce about 300 µg of Vg/day, which would allow for a maximum 1:3 nurse:open brood ratio [Omholt, Amdam 2004 op cit], which is not far from what we see during the spring turnover (see Fig. 1 in this article).
[7] Haydak, MH (1963) Age of nurse bees and brood rearing. Journal of Apicultural Research 2(2):101-103
[8] These simple and straightforward simulations were calculated based upon 100% larval survival, and the worker survivorship curve from:
Harris, JL (2010) The effect of requeening in late July on honey bee colony development on the Northern Great Plains of North America after removal from an indoor winter storage facility. Journal of Apicultural Research and Bee World 49(2): 159-169.
[9] In The Wisdom of the Hive, p.183.
[10] Pankiw, T, ZY. Huang, ML Winston, GE Robinson (1998) Queen mandibular gland pheromone influences worker honey bee (Apis mellifera L.) foraging ontogeny and juvenile hormone titers. Journal of Insect Physiology 44: 685–692.
[11] Naumann, K, ML Winston, KN Slessor (1993) Movement of honey bee queen (Apis mellifera L.) mandibular gland pheromone in populous and unpopulous colonies. Journal of Insect Behavior 6: 211–223.
[12] There could be some benefit to the swarm during its establishment, but swarms (or package bees) generally seem to be able to build quite quickly despite the 35-day mean longevity limitation.
[13] Preferred cavity size and populations from Seeley (1995) (below) and Winston’s The Biology of the Honey Bee, Fig. 11.1.
[14] See Fig. 4 in What’s Happening ToThe Bees? – Part 4: The Genetic Consequences of Domestication. ABJ May 2014 or at Is There A Difference Between Domestruicated And Feral Bees?
[15] Relabeled weights, from Seeley, TD (1995) The Wisdom of the Hive. Harvard University Press.
[16] Assumptions: I used Lloyd Harris’s survivorship curve, stretching it for the 70-day calcs, and started with the Ibid expected age distributions of the workers based upon the respective survivorship curves.
| Assumptions and Calculations | |
| 173J/day resting forager, (Harrison & Fewell); 25W/kg bees, broken cluster at cool temp (Southwick).
These both worked out to about 500 Wh/day/kg bees, which I converted to lbs of honey consumed/day. |
|
| 500 | Wh/day/kg bees |
| 9 | 1000 bees/kg |
| 55.56 | Wh/day/1000 bees |
| 1.16 | Wh/kcal |
| 47.77 | kcal/day/1000 bees |
| 0.03 | lb honey/day/1000 bees |
| 1.04 | Est. lb honey consumed/day/30,000 bees; this is close to Seeley’s measured 1-lb loss per day (as a confirmatory check on my estimation) |
| There were 1,688,000 extra bee days in the 70-day hive over the 35-day hive during the course of each colony dropping to the 10,000-bee population. This works out to 58 lbs of honey consumed. | |
Southwick, EE (1982) Metabolic energy of intact honey bee colonies. Comp Biochem Physiol 71A: 277-281.
Harrison, JF & JH Fewell (2002) Environmental and genetic influences on flight metabolic rate in the honey bee, Apis mellifera. Comparative Biochemistry and Physiology Part A 133: 323–333.
[18] Visscher, PK and R Dukas (1997) Survivorship of foraging honey bees. Insectes soc. 44: 1 – 5.
[19] Yan, H, et al (2015) DNA methylation in social insects: how epigenetics can control behavior and longevity. Annu. Rev. Entomol. 60: 435–52.
[20] Scofield HN, Mattila HR (2015) Honey bee workers that are pollen stressed as larvae become poor foragers and waggle dancers as adults. PLoS ONE 10(4): e0121731. doi:10.1371/journal. pone.0121731
[21] Karsai,I and T Schmickl (2011). Regulation of task partitioning by a “common stomach”: a model of nest construction in social wasps. Behavior Ecology 22: 819-830.
[22] With different effects on the workers from the pheromones of either young or old brood.
[23] Maisonnasse A, Lenoir J-C, Beslay D, Crauser D, Le Conte Y (2010) E-b-Ocimene, a volatile brood pheromone involved in social regulation in the honey bee colony (Apis mellifera). PLoS ONE 5(10): e13531. doi:10.1371/journal.pone.0013531
[24] Traynor, KS,Y Le Conte, RE Page Jr (2015) Age matters: pheromone profiles of larvae differentially influence foraging behaviour in the honeybee, Apis mellifera. Animal Behaviour 99: 1-8.
[25] Part 8– The Main Honey Flow
[26] https://scientificbeekeeping.com/the-primer-pheromones-and-managing-the-labor-pool-part-3/
[27] Note: this study was published prior to the understanding that Vg suppresses JH, so it’s not clear how Vg was involved in the findings.
Huang, Z-Y and GE Robinson (1995) Seasonal changes in juvenile hormone titers and rates of biosynthesis in honey bees. J Comp Physiol B 165: 18-28.
[28] Rueppell, O, et al (2010) Altruistic self-removal of health-compromised honey bee workers from their hive. J Evol Biol 23: 1538-1546.


