Thanks to these sponsors, you can enjoy this website without annoying popup ads!
You can show your appreciation by clicking on their banners above to go directly to their websites.
I’m writing these words in mid-April. It’s been a miserable wet spring in Northern California (Fig. 1), and we’ve been furiously trying to catch up with delayed queen rearing while trying to stay ahead of swarming. I struggled to find time to type up my next installments on nosema and pesticides, then decided to instead write about what I was currently immersed in — my selective breeding program for mite resistance.
Figure 1. If you had trouble getting queens or packages this spring, you can blame the weather in California. I snapped this photo of an almond orchard on a cold day shortly after a hard rain had knocked some of the flower petals to the ground. These surreal black-and-white manmade forests never cease to amaze me with their unusual beauty.
Selective breeding for mite resistance
The varroa/virus complex is the number one problem for most beekeepers worldwide. As a beekeeper and biologist, I feel that our industry, after 30 years with varroa, has been derelict in not yet fully adopting honey bee stocks that are naturally resistant to the mite. To help get the ball rolling, in the spring of 2017 I wrote “Bee Breeding for Dummies” [[1] ] to suggest how most any medium-to-large-scale queen producer could engage in a serious, yet simplified, low-cost selective breeding program for varroa-resistant bees. I then promised to “walk the walk” in order to determine how much it would cost, and what sort of progress I could make.
In spring of 2018 I shared my experiences in “Selective Breeding for Mite Resistance: 1000 hives, 100 hours,” [[2]] being surprised at the minimal costs involved. And now in spring of 2019, I’m sharing another progress report after two years of experience.
Full transparency: These articles are not a sales pitch — I’m not looking for new customers. Instead, what I’m trying to do is to see whether selective breeding for mite resistance is something that Joe Queenproducer can do without any scientific training or equipment, using his own stock of bees as the starting point. If I’m not successful, I’ll tell you. If on the other hand, I am successful, then there will be no excuse for every queen producer not to be breeding and selling mite-resistant queens.
The 2018 season
After heavy culling of our remaining potential breeders in the fall of 2017, the following spring we bred from the queens of only the 20-some colonies that exhibited minimal mite levels after a year without treatment (roughly 2% of our 2017 starting count of 1000 hives in the program). I was somewhat disappointed that by April, the mite counts in most of those breeders had started to climb a bit. But I had to keep in mind that those queens had been mated to a drone pool coming from almost entirely non mite-resistant stock, so I really couldn’t expect to see much improvement yet.
Anyway, for 2018 we started over 1500 new colonies with daughters from those twenty queens. Come late June, when I asked my sons for help with mite washing, they enthusiastically jumped on board, saying that the last-season’s across-the-board mite washes had actually paid for themselves in the savings from unneeded treatments, and resulted in lower varroa levels across the operation — perhaps due to our identifying and culling any hives with extremely high mite levels.
Mite assessment went even quicker this season, since following my article “Smokin’ Hot Mite Washin’,” [[3]] I had built several prototypes to come up with a portable, rechargeable motorized mite washer, which sped up our assessments immensely (Fig. 2).
Figure 2. My assistant Brooke Molina, with the portable mite washer. The washer plugs into a cigarette lighter socket to recharge between yards, and can be easily carried right into the apiary. We’ve performed nearly a thousand washes with this prototype, and absolutely love it. I plan to make it available to all as soon as I can, as it revolutionizes the concept of monitoring for varroa — taking less than a minute to get an accurate mite count. You can see a short video of it in use at [[4]].
Brooke has by now counted enough mites to last a lifetime — often needing to count 50 or more in a cup. But with last year’s potential breeders, their low mite counts allowed us to move very quickly. Zeroes get big smiles!
Practical application: What we found was that with a crew of five, we could obtain alcohol wash counts from a yard in less than a minute per hive — from truck doors opening to doors closing. And instead of being tedious work, it was actually fun to do as a crew — at each stop we all jump out and hit the ground running — each of us with specific jobs, racing against each other, joking and laughing (Figs. 3- 5). Then off to the next yard.
Figure 3. In this snapshot, Brooke caught five of us blasting through a yard for the mid-June first assessment. These colonies had been started as late nucs in April, and had nearly finished drawing the foundation in their second brood chambers. At this time point, any colony with an alcohol wash count of over 2 mites per half cup of bees, or that hasn’t grown to above yard average, didn’t make the grade. This eliminates around 90% of the colonies in each yard from further consideration as breeders. We also use this inspection to take care of any hives that need attention.
Figure 4. In order to keep track of which mite count goes with each colony, we use color-coded cups and matching hive markers (not shown). The person counting mites simply looks for the matching color marker on a sampled hive, writes down the mite count, and then puts the now-refilled cup and marker on the next hive to be tested. The system works beautifully, with minimal confusion.
Figure 5. Move fast, but don’t rush! My sons found an exceptionally productive hive last season, and suggested that I check it for mites. It was late on a cold day, so I hurriedly took a sample. To my horror, as soon as I dumped the bees into the alcohol, my eye spotted the queen’s abdomen. Unfortunately, there’s no way to save a queen once she gets immersed. The mite wash turned out to be zero, so I immediately grafted 100 daughter larvae in order to save her genetics.
Our mid-June assessment of 1500 hives in 50+ yards took us several days. Only 171 hives made the grade of having mite infestation rates of less than 1% (Fig. 6).
Figure 6. Of our starting 1500 hives, only 171 (11%) made it past the first assessment. By November, there were only 80 (4%) still in the running.
We minimize recordkeeping by simply stapling an index card on the lid of any potential breeder hive (Fig. 7).
Figure 7. I keep it as simple as possible. At the first assessment, any colony making “potential breeder” status gets a card stapled to the hive cover (it doesn’t rain during the summer where we live). We write the dates and alcohol wash count of the hive at each assessment, or immediately eliminate the colony as a potential breeder if the mite count is above what we consider to be acceptable — at which point we remove the card and treat the hive. In case you’re curious, the above hive scored zero mites at the following late March assessment (not shown).
Once we’re down to fewer than 100 potential breeders remaining, I pin a numbered stainless steel tag on the landing board and record all the previous mite counts from the index cards, so that I don’t lose track of the information over the winter (Fig. 8).
Practical application: We’re learning as we go. I want to keep the system as simple as possible, and keep paperwork to a minimum. To date I haven’t done formal tracking of bloodlines, but am planning to make an attempt to do so this season.
Figure 8. I find these off-the-shelf stamped stainless steel tags to be handy to identify potential breeder colonies, since our hives are otherwise unmarked. We record the mite counts from the index card when we apply the metal tags in fall.
Last season (2018) we didn’t seem to experience the same degree of September mite immigration that we had in 2017 — which in that year had knocked around half of our August potential breeders out of the running. In correspondence with Dr. John Kefuss, he suggested that I should have allowed some of those hives to try to recover. So in November of 2018 I retained potential breeders with infestation rates up to 3 mites/hundred bees (Fig. 9), and was glad I did.
Figure 9. Note the typical trajectory of mite increases, between the June and August assessments, even in a number of colonies tentatively labeled as potential breeders in June. Since we assess these potential breeders roughly once a month, no colonies die from mites in this program — those not making grade are immediately treated with an appropriate organic acid or thymol. The heavy black line represents the average expected progression of mite increase in a typical untreated hive in our area — generally reaching a 20% infestation rate (60+ mites in an alcohol wash) by October. The light pinkish line going up from mid-August is a colony in which the varroa count for some reason exploded. Note how few potential breeders needed to be culled after the late August assessment, despite late-season mite drift.
A visit to France
I’d long corresponded with Dr. John Kefuss — the man who coined the term “The Bond Method” for allowing nature to take its course in selecting for varroa-resistant bees. My own proposed breeding program is modeled on his “Modified Bond” method, applies the same selective pressure, but without the loss of any colonies, so I wound up being invited to present the method to the French National Association of Bee Breeders (ANERCEA). Afterwards, John took me to visit his main apiary (Fig. 10).
Figure 10. Dr. John Kefuss in his main apiary in Toulouse, France, where he invited me to participate in his “World Varroa Challenge” and earn a shiny new French penny for each mite that I could find. This was in November, so it required me to dissect brood in six hives before I could add my name to the list of notables who had earned their penny. John has clearly demonstrated how the application of strong selective pressure can result in the production of stock that requires no varroa treatments.
My spring surprise
We couldn’t resist the offered fee for almond pollination, so took most of our remaining 70 potential breeders to the orchards, where they easily made grade. Most continued to build up even during the miserably-rainy bloom period — we only consider colonies as potential breeders if they perform well. As they returned from almonds, I took mite washes, ready to dismiss those in which mites had built up since the November assessment. The colonies had begun broodrearing in earnest 10 weeks earlier — that’s over 4 varroa reproductive cycles — after a very brief winter brood break. Thus that time period would have allowed the mites to build up. But one must correctly interpret alcohol wash counts:
Practical application: This entire selection method is based upon using alcohol wash assessments to identify colonies that somehow restrict varroa reproductive increase. So it’s important to understand at every time point how the alcohol wash counts relate to the total mite population in the hive. For this, it’s helpful to use my open-access varroa model. [[5]] The model suggests that during the period from January 1 through April 1, that the mite population in a California hive would have built up 11-fold. But due to the large increase in the amount of worker and drone brood present, an alcohol wash count would only double. Take home message: Alcohol wash counts underestimate varroa buildup early in the season, but then overestimate mite population increase once the colonies begin to reduce broodrearing later in the season.
To my great surprise and delight, as of mid-April, 25 of the returned potential breeders exhibited mite wash counts of zero or only a single mite (well less than a 1% infestation rate). Incredulous, I made a point of inspecting the drone brood of these hives to see whether there were mites in hiding. As you can see in Figure 11 (from a representative breeder hive), no mites were to be seen.
Figure 11. Note the absence of mites in the drone brood of this breeder colony after return from almonds in early April. I checked each breeder colony’s drone brood to confirm the results of the alcohol wash.
Practical application: A mite count of zero does not mean that a colony at this time of year is devoid of mites, but it does indicate that the mite infestation rate has certainly not increased to any extent over the course of a full year without treatment.
Selection vs. Bottlenecking
Any selective breeding program consists of bottlenecking the genetic diversity of the breeding population — in this case by applying strong selective pressure for the trait of mite resistance. But in the case of honey bees, we must always keep in mind that one needs to maintain enough diversity in the sex alleles in order to ensure viable worker brood. Thus, I try to strike a balance by breeding off of at least 25 different queen mothers each season. Below are the track records for my 28 breeders chosen for this spring (Fig. 12).
Figure 12. Mite infestation rates over time of the colonies whose queens who made the grade for 2019. Note that of the 28 selected breeders for this season, none exhibited infestation rates of over 1% after a year without treatment. The red dashed line indicates the 3-mites-per-100-bees infestation rate that might have been cause for concern in the fall. The few that I chose to keep despite their showing higher counts at that time were kept in the running because they were exceptionally gentle, productive, and came from high-mite yards. But perhaps most impressive, they then brought those counts back down without help.
I’m also going to graft from a breeder now in her second year without treatment. The incredibly exciting thing for me is that 13 of those breeders never reached a mite infestation rate above 1% over the entire course of a year — again without any treatment. And every breeder had mothered a gentle and productive colony.
Practical application: When I see such lovely colonies handling varroa on their own, it gives me goosebumps!
Choosing the breeders
Despite exhibiting low mite counts, no queens were chosen as breeders unless they went to, and returned from, almonds strong and healthy. I entered their returning mite wash counts into my spreadsheet, along with notes from last summer, and then used the sort function to rank them by the combined total number of mites from those five assessments. I then assigned each colony an additional letter name (since we use letter names to keep track of which mothers we’ve grafted from); this also now ranks the breeders alphabetically by preference (Fig. 13).
Figure 13. I prioritized Queen A, whose colony in its second year continued to control the mite. Those that made an exceptional amount of honey last summer were labelled “productive.” Note that colony #54 had a combined total of only 2 mites in the 5 alcohol washes taken over the course of a year. Since I always worry about a breeder queen swarming prior to grafting, I graft from the most desirable first (hence my ranking of them alphabetically).
The big question ― heritability
Hey, colony #54 is a gem, but is that apparent trait of mite resistance actually heritable via grafting from their queen? I have no idea how these colonies managed to keep their mites in check, but if the trait isn’t heritable via her daughters, a selective breeding program would be a waste of time.
Here’s the problem: The trait of “mite resistance” is something that we observe, not a definition of how the bees do it. As an analogy, we may observe that a basketball team appears to express the trait of consistently winning games. It could be pure luck, or something inherent in the team. Different teams may be winners for different reasons — it may be due to a couple of star players, or the team’s offense, their defense, their ability to rack up three-point shots, or the coach’s skill at choosing which players work well together on the court.
Similarly, there are any number of ways that a colony could gain resistance to varroa, [[6]] and as with basketball, it may be something in common for every member of the team, or due to different members having specific abilities that all come together in the hive.
So when I identify a colony exhibiting consistently low mite levels, that may be due to all the workers in the hive expressing the same genes, or it may be due to some lucky combination of sister patrilines (each fathered by different drones that the queen had mated with) all working together.
In order to develop a bloodline of bees that can consistently control the mite, the entire suite of necessary alleles (forms of each gene involved) must be carried by each breeder queen, and perhaps most of the drones that she mates with (this is likely why so many beekeepers wind up being disappointed when they’ve open mated the daughters of instrumentally-inseminated resistant queens).
Practical application: It’s not the queen who fights the mite — it’s a team effort of the various patrilines of workers in the hive. Thus, although a colony may exhibit mite resistance, that doesn’t necessarily mean that new colonies founded by daughters of the resistant colony’s queen will carry all the critical alleles.
In my own program, going into its second season of mating daughters solely from the queens of colonies exhibiting apparent mite resistance, it’s difficult for me to resist a glimmer of hope:
During breeder selection in 2017, none of them exhibited mite counts of zero the next spring.
This year, at least 12 did.
So it would be easy for me to think that I’m making progress. But I must temper my excitement, since the other 98% of the hives in our operation still required mite treatments. The big question is, is the apparent [[7]] mite resistance of my selected breeders heritable? Other breeding programs have demonstrated that it can be, so I have hope, but I don’t expect a miracle.
Some things to keep in mind are that not all those selected colonies necessarily used the same mechanisms to control the mite, meaning that when their sisters (yes, next season’s queens are still daughters of their mothers, and hence sisters of the workers) cross mate, those mechanisms may conflict. And even assuming that their apparent resistance is genetically determined, it’s likely that there would be a number of protein-coding or regulatory genes involved, and if so, it then makes a huge difference whether those genes exist on the same chromosomes, and furthermore, whether they are close enough on the DNA strand to be “linked.” Not only that, are those alleles dominant or recessive (in which case the drones would need to carry the same alleles)? And I’m also fighting the high recombination rate of the bee genome during reproduction, which keeps mixing the genes up. [[8]]
Practical application: Apologies for the scientific jargon, but I just want to assure you that I’m not approaching this blindly.
The importance of the drone pool
Over the years, many of us breeders have brought in instrumentally-inseminated mite-resistant queens, only to watch that desirable trait disappear in subsequent generations. So you have every reason to ask, Randy, (1) why aren’t you using instrumental insemination, and (2) what makes you think that you can exercise control of the open matings of the daughters of your selected queens? My answer to the first question is: because I want to keep it as simple and inexpensive as possible. As far as the control of matings, luckily, some USDA researchers answered that question back in the 1990s by cleverly allowing daughters of double-recessive cordovan queens to open mate in or around commercial queen breeding apiaries. [[9]] Their conclusion was:
We consider 90-95% to be a realistic level of mating control that most queen producers will be able to attain without substantially modifying existing practices.
Practical application: I have the greatest respect for our commercial queen producers — they serve our industry well. But if you’re going to seriously breed for mite resistance, you need to go all in.
The point is, that in order to control matings, you must restrict the production of drones in all nearby colonies solely to daughters of selected breeders (similar to how the producers of the hybrid Starline and Midnight queens did back in the day). In the case of breeding for mite resistance, that means that every queen in the operation must come from the mother of a colony exhibiting resistance.
Practical application: 90% of control of matings is good enough for me. We replace every one of the queens in our operation with daughters from selected breeders each season, and run a drone frame in our hives when we go to almonds, resulting in colonies bursting with drones. We thus flood our mating yards year after year with our own “selected” drones. My hope is that this will result in balancing our inbreeding for mite resistance vs. the maintenance of sufficient genetic diversity in my breeding population.
The above said, I was curious as to just how much progress I might expect from open mating.
A Primer on Bee Genetics
The production of eggs and sperm during meiosis involves a mixing of alleles from the father with those of the mother, in order to create novel genetic combinations. In honey bees, the mixing up of genetics each generation is amplified, due to the multiple matings by the queen, and the bees’ unusually high rate of genetic recombination. Such genetic blending helps to allow honey bees to rapidly evolve, but may make it tough to breed for specific traits.
Half of any queen or worker’s alleles (on average) come from her mother, and half from only one of the many drones that her mother had mated with. And I have no idea which drones carried the alleles critical for mite resistance. Thus, without incorporating single-drone instrumental insemination into a breeding program, it may be more difficult to select for the critical alleles involved. I’m fully aware of this, but choose to stick with open mating since that’s what Joe Queenproducer can easily do.
That said, his/her only control of the genetics of the drone pool is by making sure that all drones in that pool come from daughter queens produced in the previous year solely from larvae of queen mothers of colonies exhibiting strong mite resistance — knowing full well that not all of those drones will necessarily carry alleles for mite resistance.
Practical application: Each season’s virgin queens will supply all the genetics for the next season’s drones — I’m counting on this, rather than using instrumental insemination. But since queens come from fertilized eggs, half the genes of the daughters of a breeder queen come from their mother, and half from one of the drones that their mother had mated with (the new drone’s grandfathers) — but no telling whether that particular drone carried genes for mite resistance. Thus, even with complete drone flooding, the best that one can hope for is to shift the genes responsible for mite resistance in the drone population by less than 50% each season.
In order to illustrate this visually, I created the diagram below (Fig. 14) in which, I’m selecting a small percentage of my stock to be breeders each season (surprisingly, that percentage doesn’t affect the proportional progression illustrated). The red area of each pie represents alleles from unselectedcolonies; the blue area represents alleles carried by the selected queens (although not necessarily alleles for resistance). The chart assumes a best-case scenario of a full 50% shift of the genetics of the drone population each season — but that shift does not necessarily represent the degree of shift of the genes responsible for mite resistance.
Figure 14. Even though I selected the queens from our most mite-resistant colonies as breeders last season, their daughters mated with a drone pool coming from 98% unselected stock. The virgins that we produce this season will mate with a drone pool in which half the genes will now trace back to those from selected queens.
Practical application: The above illustration represents the absolute best case scenario for genetic progress, since I have no idea as to how many genes are involved in the mite resistance of my breeders, nor whether all the breeder colonies depended upon the same genes. Thus I certainly don’t expect to see much improvement in mite resistance for at least another couple of years, since it will take a while to fully shift the genetics of the drone pool. But seeing how many apparently-resistant breeders I have this spring, I can’t help but being “guardedly optimistic” about what I hope to observe in 2020.
Bottom line
Can breeding for mite resistance be a simple and straightforward process that does not require scientific training or loss of colonies, and only minimal expense? In theory, yes — indeed, this sort of “directed evolution” was how all breeds of vegetables, fruits, and livestock were created prior to Gregor Mendel’s explanation of inheritance. What I’m doing is to see the extent of progress that a thousand-hive operation can expect to make in a reasonable amount of years.
Practical application: In order to prevent loss of sex alleles while applying strong selective pressure, it will likely take having at least 1000 hives in a breeding program. This is why I’m targeting commercial queen producers.
Note to hobby beekeepers: I say this to make clear that beekeeper Jane or Joe Treatment-Free is dreaming if she or he think that allowing their colonies to die from lack of mite management is going to improve honey bee genetics overall. If they want to improve honey bee genetics, they’d do best to treat their hives to keep them healthy, but put pressure on whoever they purchase queens from to seriously select for mite resistance.
Note to commercial queen producers: I’m making no promises, but am merely sharing what I learn as I determine whether my proposed simplified selective breeding program is worth pursuing. If it proves out, it’s so simple and cost effective that it will eliminate all excuses for you not to be seriously breeding for resistance. If it doesn’t work, rest assured I’ll let you know as well.
I gotta tell you though, seeing these strong, gentle, productive hives that are able to maintain minimal mite levels over the course of an entire year of near-continuous broodrearing in California has got my crew danged excited about the possibility of a future in which we would no longer need to worry about varroa and its associated viruses.
Acknowledgements
Thanks to my hosts in France ― John Kefuss, Yves Le Conte, Benjamin Poirot, and Gabrielle Soland, with whom I discussed this proposed breeding program. Also Albert Robertson (Saskatraz queens), and all the beekeepers and queen producers supporting me in performing this project. And as always thanks to Peter Borst for his library assistance.
[7] I’m being careful to qualify what “appears” to be resistance. My tracking of mite levels over the course of a full year tends to support the assumption that the low mite levels in the breeders are not simply flukes.
[8] “High recombination rates appear to have evolved independently in several eusocial insects, leading to the hypothesis of its association with the evolution of eusociality, particularly caste specialization. For example, high recombination can allow simultaneous positive selection of beneficial mutations and negative selection of deleterious mutations by decoupling the effects of linked alleles.” From:
Dogantzis KA, & A Zayed (2018) Recent advances in population and quantitative genomics of honey bees, Current Opinion in Insect Science https://doi.org/10.1016/j.cois.2018.11.010
[9] Hellmich, R & G Weller (1990) Preparing for Africanized honey bees: Evaluating control in mating apiaries. ABJ 130(8): 537-542.
Hellmich, R, et al (1993) Evaluating mating control of honey bee queens in an Africanized area of Guatemala. ABJ March 207-211.
It’s “Common Knowledge” that Nosema Causes Dysentery…
But is it Actually True?
First published in ABJ May 2019
Randy Oliver
ScientificBeekeeping.com
Yesterday I inspected a group of 40 hives returning from almond pollination. All were strong and healthy, other than two unusual deadouts, which no longer had any bees on the combs, and only a scattering of a few dead workers on the bottom boards. One hive had clear signs of dysentery on the top bars; the other didn’t. Upon further inspection, there were small arcs of apparently-healthy sealed brood remaining in both hives, and surprisingly, fresh eggs in the cells below that brood. What could be the cause of these unusual signs?
From my experience, the field signs above point the finger at nosemosis — as described by The World Organisation for Animal Health (OIE) [[1]]:
In a typical case of a colony being depleted because of a Nosema infection, the queen can be observed surrounded by a few bees, confusedly attending to brood that is already sealed.
Exactly as in the photo below, which I took during the CCD epidemic in the early 2000s.
Figure 1. Ten years ago, many beekeepers reported unexplained Colony Collapse Disorder, not associated with varroa. Colonies would quickly dwindle to a “silver dollar sized patch of bees and a queen,” as in this photo. Often times there would be patches of healthy brood remaining after the bees disappeared. I strongly suspect that a large part of the CCD epidemic was a result of the invasive wave of Nosema ceranae, as a part of a “perfect storm” that included evolving viruses, failure of varroacides, and evolving strains of European Foulbrood.
Practical application: Colonies that exhibit healthy brood during cool springtime weather, yet fail to build up, or in which the adult population appears to be declining, may well be suffering from nosemosis.
Nowadays, most of what is now termed “Colony Collapse” is caused by varroa and Deformed Wing Virus (DWV), and can be easily differentiated from collapse due to nosema by the telltale signs of brood dying from the virus, as well as the presence of mite fecal deposits on the ceilings of the cells (Fig. 2).
Figure 2. A typical varroa/DWV deadout, which typically occurs late in the season. Some cells contain dead pupae, or fully-developed adults that were unable to emerge. Note the distinctive traces left by varroa — the white fecal deposits at the top of the cells.
Practical application: Before you blame the collapse of your hive on something else, first pull a central brood frame out of the deadout, and check for the signs above.
But I still occasionally see colonies collapsing from nosema in early springtime. So how about my two deadouts — could nosema have been the cause? And why would only one hive exhibit dysentery? Again from OIE:
In some acute cases [of nosemosis], brown faecal marks are seen on the comb and the front of the hive, with sick or dead bees in the vicinity of the hive. However, the majority of colonies show no obvious signs of infection, even when the disease is sufficient to cause significant losses in honey production and pollination efficiency.
Practical application: So how could I tell whether nosema was the culprit? There’s only one way to tell whether a colony is infected by nosema ― I’ll put it in large type:
The only way for a beekeeper to diagnose nosema infection is by microscopy.
And sure enough, when I crushed dead bee samples from each of the bottom boards, Nosema ceranae spores were clearly evident and in abundance (Fig. 3).
Figure 3. Nosema spores ― the numerous small glowing elongated ovals — from a diluted sample of crushed bees viewed at 400x. The sample from the hive with dysentery looked identical to the one without.
Practical application: Get a microscope, or encourage your local club to purchase one. My favorite for viewing nosema is the Omano 36 [[2]], but even a cheap secondhand ‘scope will do.
So, yes to nosema, but no to having a correlation with dysentery. In fact, I’ve yet to find a single study that has ever demonstrated that nosema ― whether N. apis or N. ceranae — is responsible for causing dysentery.
A request and challenge: If you can find any study that has experimentally confirmed that nosema causes dysentery, please forward it to me.
What is Dysentery?
We’ve all seen it ― dysentery on the front of a hive. Clearly those bees “really had to go” and let loose the moment they took off (Fig. 4).
Figure 4. The telltale signs of dysentery. Even worse is when it’s all over the top bars inside the hive, since it could then quickly spread any gut pathogens or toxic substances throughout the cluster. Photo credit: Monique Vescia
Honey bee workers are remarkably fastidious about hygiene, and do everything possible to avoid defecating within the hive. Foragers wait until they’re on the wing, whereas non-foraging “house bees” take “cleansing flights” when necessary.
Many of us have had to apologize to our neighbors about the orange polka dots on their cars or clotheslines. Although those drops of bee poo are normally a mere annoyance, back during the Viet Nam War, it became an international issue, when accusations were leveled about purported chemical warfare — the evidence being spots of “yellow rain” dotting leaves [[3]]. Such “rain” would have been immediately recognized by any beekeeper who’s enjoyed the experience of walking in an apiary when the sun comes out after the bees had been confined by weather to their hives for several days.
During periods of forced confinement, honey bees have a remarkable ability to “hold it” in order to avoid soiling their nest (for months if necessary). A bee’s rectum has the ability to distend to the point that it nearly fills the abdomen (Fig. 5).
Practical application: Dysentery is an indication that a colony has a serious problem ― one that sometimes leads to the death of the hive, but not necessarily from nosema. I will later return to the subject of the causes of dysentery.
So Why Does Everyone Think That Nosema Causes Dysentery?
Here’s where we get into one of my pet peeves about honey bee popular books, as well as scientific papers ― people tend to repeat something they’ve heard just because it seems to make sense — but without first actually checking the facts.
I see it in scientific paper after paper. The author says that dysentery is a “sign of nosema” and throws in a citation to support that claim. I’ve checked dozens of those citations, and have yet to find a single one that actually refers to any study that actually demonstrated that nosema infection induced dysentery. Keep in mind that nosema infects the cells that line the bee’s midgut; the “environmental spores” that are found in the hindgut have never been shown to infect or irritate the lining of the rectum. And if nosema did indeed cause lack of hygienic control, you’d expect to see the fecal contents leaking out as little dribbles, not in the huge streaks from a rectum able to be kept full to bursting.
Unfortunately, many authors don’t bother to check, and something that someone once said gets repeated over and over again until it becomes “common knowledge.”
Practical application: This is not the only “common knowledge” myth in the bee literature that possibly needs to be debunked ― I’ll be addressing others in the future.
The Sad Part
The really sad thing about this misconception is that researchers and textbook authors have no excuse for repeating such an assumption, since the lack of nosema-induced dysentery was well established by 1922, and the cause of dysentery by 1935. But I’ve rarely seen any of the many recent papers on nosema cite those nicely-aged, but scientifically accurate studies.
Practical application: Not every author or researcher does their homework. I’ve learned over the years to double check the supportive citations in scientific papers myself, a process that often leads me down a string of misinterpretations (and sometimes to different conclusions than reached by the authors) [[5]]. I’m often appalled by the amount of scientific sloppiness allowed to get by the peer reviewers of papers.
Nosema Apparently Does Not Cause Dysentery
I discovered this when I began deep research into N. ceranae in 2006, and found that most everything that we needed to know about nosema had been clearly explained in a forgotten USDA bulletin published in 1919 [[6]]. I love to read these old studies, and am often impressed by the scientific diligence of the government-funded authors — in this case a G.F. White spent 9 years experimenting with Nosema apis in every way. White’s 58-page summary, in my opinion, still stands as perhaps the most informative study on the parasite, and the disease it may cause, yet published in the English language.
Practical application: One thing that I found was that N. ceranae was not all that different from N. apis. Once the invasive wave of N. ceranae passed through North America in the 2000s, the main differences that I now see between the two species is that ceranae may crop up under certain conditions during warm weather, its spores are far less cold tolerant, and its spore counts may be much higher (although they do not cause a whitening of the hindgut, as with N. apis). Both species are common parasites of the midgut in springtime (with ceranae now being far more prevalent than apis), generally appear to be rather harmless; but may cause disease in stressed colonies.
Allow me to quote Dr. White:
“Care should be observed that Nosema-disease is not confused with dysentery…In fact the two disorders are very different and should be considered, for the present at least, as having no direct relation to each other. As both conditions are widely distributed and occur most frequently in the spring of the year. It is to be expected that not infrequently both of them may be encountered together in the same colony.”
The start of the “association” between nosema and dysentery is perhaps explained by famed bee pathologist Dr. Leslie Bailey [[7]]:
Infection by Nosema apis is commonly believed to make honeybees (Apis mellifera) void faeces in and on the colony instead of away in flight — a condition known as ‘dysentery’ by beekeepers. Evidence that seemed to support this was provided by Lotmar (1951), who found more faeces accumulated in caged infected bees than in healthy ones. No field experiments have shown this effect…
Dysentery was not caused primarily by N. apis, because it occurred whether colonies were severely infected or not…Most of the colonies that survived managed to clear the faeces away; the few still with combs contaminated with faeces were among those with more than 25% of bees infected… If infection is spread by dysentery, this is not unexpected, since infection would spread most in colonies that suffered most dysentery; but if infection causes dysentery a more striking correlation between dysentery and severe infection would be expected than actually occurred.
Practical application: Let me make clear that should a bee already infected with nosema happen to defecate within the hive due to some other gut issue, it will certainly transmit nosema spores to the other bees that clean up the mess. But the dysentery was apparently the result of something other than nosema itself.
Part of the apparent confusion between nosema and dysentery may be due to the fact that they both commonly occur in early spring. In my own apiaries, dysentery comes and goes in the springtime, but after checking numerous dysentery samples (from my own apiaries and numerous samples from others) under the microscope, I have yet to notice any correlation between nosema and dysentery.
Biological evaluation: One could reasonably expect a gut parasite to induce dysentery in its host, as a means of transmission of its spores. But if nosema did indeed induce dysentery, every infected colony would quickly be overwhelmed by spore transmission, and die before spring. This we do not see. Could it be that both species of nosema are actually rather benign parasites, which under “normal” circumstances do not cause appreciable harm to the colony? That is, unless the colony suffers from dysentery due to some other reason.
In France, the impact upon colonies from N. ceranae has been referred to as “dry nosemosis,” since it is clear that the parasite doesn’t cause dysentery. Perhaps we should now question whether N. apis ever did either.
next
In my next two articles I’ll discuss our current state of knowledge about Nosema ceranae ― its seasonality (and the causes for that seasonality), its effects upon colony performance (or lack thereof), and the best ways to monitor for it.
And then I’ll follow with an article about the potential causes of dysentery in bees.
The Pesticide Situation Part 5: Reinventing Nature
Randy Oliver
ScientificBeekeeping.com
First published in ABJ May 2019
In their never-ending battle against herbivores and parasites, plants have for eons been synthesizing toxic chemicals, collaborating with microbes, or assisting predacious insects to deter or kill pests. Now that we’ve perhaps begun getting over our synthetic pesticide hubris, and are realizing that we’re running out of new chemicals, we can learn from nature itself.
Reinventing Nature
Keep in mind that plants have successfully fought pests since long before we humans invented synthetic pesticides. We’re now starting to revisit how they do this, with the goal of developing more sustainable pest control products.
Biopesticides (aka “biologicals”)
Long favored by organic farmers, pesticides that fall into the above category tend to be pollinator- and eco-friendly, and perhaps even more importantly, are desirable to consumers — especially the Millennials who are now starting to determine market demand. Biopesticides have a lot going for them [[1]]:
They are inherently less harmful to the environment,
They are typically less hazardous to the applicator,
They are designed to target only one or a few target species,
They are often effective at low application rates, and some may self-propagate,
They rapidly biodegrade,
They are preferred by consumers,
And they work well in Integrated Pest Management strategies.
Biopesticides currently account for about 5% of the pesticide market, but that proportion is expected to increase substantially. As one of the leaders in this field, Pam Marrone, explains [[2]]:
Governmental agencies are further pressuring growers by restricting or banning certain forms of conventional chemical pesticide usage… At the same time, a number of supermarket chains and food processors, key purchasers of specialty fruits, nuts and vegetables, are imposing synthetic chemical residue restrictions, limiting options available to growers close to harvest.
For example, the European Union passed legislation…that restricted and removed hundreds of pesticide active ingredients. This was followed by passage of the Sustainable Use Directive, which legislated reductions in chemical pesticides and increasing use of integrated pest management and alternatives such as biopesticides. Several countries passed programs to accelerate biologicals, for example, in Brazil and China, allowing biopesticides to get to the market more quickly and with less capital than chemicals.
Practical application: A switch to biopesticides could be very good for pollinators. And some can be best applied by bees themselves [[3]]. All projections are that biopesticides are on track to help us to manage resistant pests in a more sustainable and eco-friendly manner (Fig. 1).
Figure 1. Biopesticides are a hot topic worldwide. Although the definition varies, biopesticides can include fungi, bacteria, viruses, parasites and predators, plant extracts and oils, minerals [[4]], pheromones, hormones, RNAi or genetic engineering.
Resistant cultivars
Since plants can’t run away from predators, they fight back with chemistry. There has been a continual evolutionary chemical warfare between them and insects. Plants produce a dizzying array of chemical repellents, anti-feedants, and toxic phytochemicals in order to ward off insects. For example, some traditional maize strains in Mexico produce pollen that acts as an herbicide against competing weeds, and our favorite herbs and spices produce those aromatic molecules to deter herbivores from feeding upon them (which is why bees don’t like having essential oils dumped into their hives). But as we’ve selected strains of plants in order to make them more desirable as food for us humans, this may come at cost — if those cultivars also lose some of their protection against pests.
Practical application: The ancestors from which we’ve bred many of our favorite food crops were often poisonous or irritating to humans. Although we’ve bred out the most dangerous plant chemicals, our favorite foods still contain thousands of completely “natural and organic” phytotoxins, as elucidated by Gold [[5]].
Of all dietary pesticides that humans eat, 99.99% are natural: these are chemicals produced by plants to defend themselves against fungi, insects, and other animal predators … Each plant produces a different array of such chemicals. On average, the Western diet includes roughly 5,000 to 10,000 different natural pesticides and their break-down products. Americans eat about 1,500 mg of natural pesticides per person per day, which is about 10,000 times more than they consume of synthetic pesticide residues… Even though only a small proportion of natural pesticides has been tested for carcinogenicity, half of those tested…have been found to be carcinogenic in rodents; naturally occurring pesticides that are rodent carcinogens are ubiquitous in fruits, vegetables, herbs, and spices.
Good news: Plants developed flowers, nectar, and nutritious pollen in order to attract pollinators, but when breeding for productivity, we sometimes inadvertently bred some of those traits out of certain cultivars. Some good news from Tennessee Department of Agriculture Apiary Inspector William R. Frazier Jr is that Pioneer is phasing out its soybean cultivar 47T36, which provides scant nectar for bees, and replacing it with P46A57BX, which is far more bee attractive.
Before the invention of manmade insecticides, traditional farmers would select for cultivars that repelled pests on their own. Many of those resistant cultivars still exist. Unfortunately, as pointed out by Adaskaveg [[6]], it may take years to incorporate those genetics by conventional breeding into perennial tree and vine crops, plus “crop characteristics selected by the breeder to meet consumer demands often result in higher disease susceptibility of the crop.”
Today’s plant breeders are revisiting this natural pest resistance, both by reincorporating genes for pest resistance from ancestral varieties (Fig. 2), or by introducing species-specific toxins into crops, such as genetically-engineered Bt “Cry” (for “crystalline”) proteins into corn and cotton. The huge advantage of this technology is the specificity of the toxic action — in the case of the Cry proteins, they only target insects that actually take a bite out of the plant (as opposed to having to be needlessly sprayed all over the field), and can be specific for beetles and caterpillars, while remaining relatively harmless to pollinators and other beneficial insects [[7]].
Practical application: The adoption of Bt crops has greatly reduced the application of pesticides on U.S. cropland, with little apparent environmental cost. On the other hand, the widespread use of Roundup Ready® crops has reduced the erosion and fuel associated with weed control by tillage, and helped to increase carbon sequestration in the soil, but at the environmental cost of increased use of glyphosate herbicide and the reduction of beneficial insects that previously fed upon those weed species.
Figure 2. Some years ago I worked with an Israeli startup seeking to use RNAi to make tomato and potato plants resistant to a devastating disease carried by the psyllid insects above (in a leaf-inoculation cage of my own design). In my lab I found that some tomato cultivars were already resistant to the disease. Nowadays, breeders could use precision CRISPR gene editing to transfer those genes for resistance to currently-susceptible cultivars, thus eliminating the need for weekly spraying of neonicotinoids to control the psyllid.
Practical application: There is huge potential in “precision breeding” of plants via genetic engineering — by either reintroducing natural genes for pest resistance, or by creating novel ways for crops to defend themselves in an environmentally-friendly manner. The sooner we drop the unfounded negative stigma of “GMO,” the more quickly plant breeders can develop cultivars that are innately resistant to pests.
Intensive Agriculture
One commonly-proposed answer to The Pesticide Situation is to shift to “organic” agricultural practices. What’s not to love about “organic” farming? But what we need to do is to differentiate between the term “organic” as used as a marketing term — riding on the common misconception that it implies being free of pesticides, as compared to the concept of agro-ecological farming — focused upon maximum sustainability and minimal environmental impact. Although there is considerable overlap between the two, the arbitrary restrictions for “organic” certification don’t allow some options that may be better for biodiversity and the environment in the Big Picture. Until we get the growth and demands of our growing human population under control, agriculture’s most promising course of action appears to be to embrace a combination of high-tech, high-intensity farming, in order to spare the conversion of our threatened wildlands to cropland.
Practical application: I’ve been a booster for organic farming for decades. But in recent years I feel that “organic” has lost its way, and is now more caught up in being “anti” this or that, rather than being “pro” feeding the human population with the least environmental impact.
Innovations in Mechanical Technology
One of the biggest challenges in agriculture is the amount of labor involved. Humans soon learned how to harness animals to drag the plow, and nowadays a single diesel-fueled tractor can have the power of hundreds of horses (thus making larger farms more efficient). California agriculture is suffering from a shortage of legal residents who want to work on farms, and as a result is in a race to convert to next-generation automation and robotics. This will likely be good news for beekeepers, since robotic pesticide applications can be more precise, and applied at night. Another exciting development is robotic weeders, which could greatly reduce or even eliminate the need for herbicide applications [watch a video[8] ]. This could really help pollinators, since as farmers are forced to switch from glyphosate (Roundup®) to dicamba herbicide (due to resistance issues), the drift of this chemical eliminates the flowering weeds around field margins.
Practical application: Robotic insect and weed control could make cropland and field margins more pollinator-friendly by reducing both insecticide and herbicide application and drift.
I’d now like to introduce two new buzzwords in agriculture:
Agroecology and “regenerative agriculture”
Farming practices are often divided into two extremes — conventional and “organic” — a marketing term allowed by the USDA. Growers may receive a premium for products that meet the standards for “certified organic,” but that premium is beginning to fade. In reality, there is a large gray area between the two extremes, with many farmers being very much concerned about sustainability and being environmentally responsible, but not willing to pursue the arbitrary restrictions of organic certification. The question then is, can such environmentally-conscious farmers still be financially rewarded even if they don’t meet the organic certification standards? According to a recent study by LaCanne and Lundgren, it appears to be so (Fig. 3):
Figure 3. This chart, from the LaCanne and Lundgren study [[9]] determined the overall profitabilityof growing corn in the Northern Plains under two different types of farming practices. Not shown here is that insect pest populations were more than 10-fold higher on the insecticide-treated farms than on the insecticide-free regenerative farms.
“Despite having lower grain yields, the regenerative system was nearly twice as profitable as the conventional corn farms. [That] relative profitability…was driven by the high seed and fertilizer costs that conventional farms incurred …, and the higher revenue generated from grain and other products produced (e.g., meat production) on the regenerative corn fields.”
It’s not that farmers can’t continue to supply our food needs, but that they can do it in a more environmentally-friendly manner. Australian conservation scientist Dr. Andrew Balmford has authored several papers on integrating eco-friendly practices into high-intensity commercial agriculture [[10]].
All that information does us little good unless it gets to the farmers themselves. Our agricultural extension services are realizing that they haven’t been keeping up with the times. A recent article in California Agriculture [[11]] points out that ag extension agents need to start adopting a “from the ground up” approach for informing growers through the use of social media and demonstration projects.
Practical application: It’s easy to complain, but more meaningful to offer solutions. If family farmers don’t break even, they may go out of business, and their land may be bought up by larger farms (due to such consolidation, the median acreage of farms today is over 1200 acres). Contrary to the example of the E.U. imposing politically-motivated restrictions top-down, it may be better to offer bottom-up demonstration projects to show how to practice profitable farming with minimal pesticide use.
To that end, Dr. Jon Lundgren (coauthor of the study above) quit the USDA in order to establish Blue Dasher Farm ― a Research and Demonstration Farm in Regenerative Agriculture, in South Dakota (Fig. 4). Jon works closely with a large commercial beekeeper, and one of his missions is to demonstrate how to practice bee-friendly farming. He is also involved in projects in California and other states. I applaud Jon for his efforts to demonstrate in real life how farmers can improve their practices — he deserves our strong support.
Figure 4. A demonstration day at Blue Dasher Farm. Similar to beekeepers, growers tend to copy the practices of other successful farmers. Demonstration farms are perhaps the best way to show them how to improve their practices.
Action item: Put your money where your mouth is. You can financially support Dr. Lundgren’s demonstration projects at https://www.ecdysis.bio/take-action/ Jon is walking the walk, and if he can show other Midwestern growers how they can profitably improve their practices to the benefit of bees and the environment, others will copy him. Disclosure: I’ve donated substantially to Blue Dasher Farm.
Resistance to change in a changing world
I feel for our farmers — they are being forced to rapidly adapt to a changing world. And many of those adaptations are not of their own choosing. There is a fundamental human objection to having the will of others imposed upon us. Farmers and landowners can be expected to resist change, unless they feel that the benefits and rewards for making the change are seen as adequate for the trouble involved. Luckily for us, one of those benefits is landowners’ common desire to be good stewards of their land for future generations (as well as not inadvertently poisoning their families). We commercial beekeepers are part of the agricultural community, and will do best if we learn to present our suggestions in a productive and positive manner.
Practical application: We beekeepers will have far greater success in getting landowners and growers to work with us if we better understand the reasons why any of us fight having to change how we do things. Our industry would do well to familiarize ourselves with what best works to overcome others’ kneejerk resistance to change. I’ve found Torben Rick’s Top 12 Reasons Why People Resist Change to be enlightening and helpful — take a look for yourself [[12]].
To be continued…
Notes and citations
[1] Gupta, S & AK Dikshit (2010) Biopesticides: An eco-friendly approach for pest control. Journal of Biopesticides 3: 186-188.
[4]According to the EPA, Biopesticides are certain types of pesticides derived from such natural materials as animals, plants, bacteria, and certain minerals. For example, canola oil and baking soda have pesticidal applications and are considered biopesticides. Kaolin clay is also commonly used. https://www.epa.gov/ingredients-used-pesticide-products/what-are-biopesticides
[10] Balmford, A, et al (2018) The environmental costs and benefits of high-yield farming. Nature Sustainability 1: 477–485. This paper is open access, with a number of useful citations.
[11] Lubell, M & N McRoberts (2018) Closing the extension gap: Information and communication technology in sustainable agriculture. Calif. Agri 72(4): 236-242.
Pesticide Resistance and Changes in Farming Practices
Randy Oliver
ScientificBeekeeping.com
First Published in ABJ, April 2019
Although I’ve heard some activists call for a ban on all pesticides, the hard fact is that production agriculture, whether conventional or organic, will for the foreseeable future depend upon the application of plant protection products to control invertebrate pests, fungal diseases, and weeds.
Unfortunately, some crop pests gain a foothold during exactly that time when honey bees have interest in foraging upon that crop — while the plants are in bloom ― thus resulting in intimate exposure of the unfortunate foragers to pesticides and their adjuvants. Notably, when it rains during almond bloom in California, most growers spray a variety of fungicides and adjuvants on the bloom — thus exposing the majority of hives in the U.S. to such cocktails (Fig. 1).
Figure 1. We were able to move our hives out of the flooding during our heavy rains just at the beginning of bloom ― others were not so fortunate. Under these damp conditions, we know that our bees will be exposed to fungicides. I’ll return to determining the effects of such spraying upon our hives later in this article.
Another example are sunflowers, which, being native to North America, come with a number of insect pests that target the flowers, making the best time to control them being exactly when bees are working the bloom. And then there are other major crops: The pollen and/or nectar of most corn, soy, canola, and perhaps vine crops contain traces of systemic insecticides, along with adhering fungicides. Thus, the unfortunate fact is that bees are often exposed to pesticides in the agricultural landscape.
Practical application: I’m no apologist for pesticide use, but I’m a realist. Our current “conventional” agricultural system has become overly dependent upon effective pest control products ― much more so than were traditional farms with multiple crops, rotating pasture, and livestock. Thus, the accepted system creates the need for pesticides, and lots of them. That’s not to say that growers couldn’t make agricultural land more pollinator-friendly — which is, commendably, the direction in which we’re headed.
The sheer number of different registered pesticides (nearly 900 sold in California [[1]]) makes for a potentially-toxic stew in the hive. And as brought to our attention by Chris Mullin [[2]], our bees must now not only deal with agricultural pesticides, but also with the often-substantial residues of miticides applied by beekeepers themselves (putting us in the position of the pot calling the kettle black). Mullin’s paper also pointed out the stunning diversity of pesticide residues found within hives.
One may ask, Why so many different types of pesticides? There are a number of reasons, including cost, ease of application, target specificity, lack of off-target adverse effects, residual toxicity, and especially efficacy. And the question of efficacy brings us to the subject of …
The development of Pesticide resistance
Plants have engaged in chemical warfare against their pests and predators since they first met (we humans prefer to consume plants that are relatively nontoxic to us). And the pests have continually fought back by evolving resistance to those toxic plant phytochemicals. The same thing occurs when a manmade pesticide is applied year after year. Insects, arachnids, and fungi have time and again exhibited the ability to evolve resistance to each and every pesticide thrown at them — often within a few generations (Fig. 2). Once a species evolves resistance to say, a certain pyrethroid insecticide, that may also confer some degree of immunity to that entire class of insecticides.
Figure 2. The blue curve indicates the cumulative number of arthropod species that have exhibited resistance to at least one pesticide; the red line shows the number of chemical compounds for which at least one species has evolved resistance. After Gould [[3]].
Insecticides are classified by the Insecticide Resistance Action Committee by their modes of action. Currently, there are 29 classes [[4]], plus a handful of others for which we’re not clear on the mode. Similarly, at least 15 classes of fungicides are used in agriculture. A number of these classes have by now been applied for decades ― thus allowing plenty of time for pests to develop resistance. This puts the chemical companies in a continual quest to develop new modes of action ― but they must now deal with much more stringent restrictions as to how eco-friendly any new product must be. The question then is whether we are fighting a losing battle? As elucidated by Gould:
Current evidence suggests that insect and weed evolution may outstrip our ability to replace outmoded chemicals and other control mechanisms.
Practical application: Pests are developing resistance faster than we’re coming up with new pesticides (Fig. 3). The upside of this for beekeepers is that farmers are going to be forced to adopt Integrated Pest Management, rather than “just spraying.”
Fig. 3. As it becomes more difficult and costly to identify and develop new active ingredients, the number of new pesticide registrations has been declining since the heyday of synthetic pesticide innovation in the late ‘60s. The dips after 1947, 1978, and 1988 were due to new regulatory requirements. Source: USDA [[5]].
It’s getting much more difficult to develop a new pesticide ― it’s estimated that companies must screen at least 140,000 chemicals in order to find one new, commercially acceptable, synthetic pesticide. The total cost to bring that chemical to market is now approaching $300 million [[6]]. And then the manufacturer’s salespeople must sell enough of the product to recover those costs before the patent runs out about ten years later ― hence the focus on developing chemicals that can be sold to a world market, and the strong sales pitches to farmers to buy them. Because of this, R&D expenditures on agricultural chemicals have been shifting toward development expenses and away from discovery research.
Again quoting Gould, pesticide resistance is not going to be solved solely by inventing new synthetic pesticides:
To avoid this outcome, we must address the mix of ecological, genetic, economic, and sociopolitical factors that prevent implementation of sustainable pest management practices.
Although the chemical companies will continue to screen newly-invented chemistries or “designer molecules” for possible development into pesticides, the regulatory hurdle is getting taller and taller. It appears that farmers are going to have to learn to use pesticides in a more eco-friendly and sustainable manner, rather than counting on the development of new chemistries.
Practical application: As with our use of fossil fuels, our agricultural practices are due for a paradigm shift.
The market demand for sustainability
Pesticide development and sales reflect the demand by the market ― not only the growers, but by the buyers. More and more we are going to hear the word “sustainability.” As elaborated by Whitford [[7]],
In the United States, SYSCO, Wal-Mart and other food companies are developing sustainable farming requirements of their grower-suppliers. In 2010, Wal-Mart announced its global sustainable agriculture goals to require sustainable best practices throughout its global food supply chain. These programs incorporate requirements for natural resource and energy stewardship, reductions and restrictions of chemical pesticides and fair treatment of farm workers.
Whitford also talks about trade-offs that can stop a company from pursuing registration of a new pesticide. For instance, if a promising new chemical kills the “beneficials” that normally check the buildup of that pest, the pest may rebound rapidly after an application of the chemical, and thus screw up overall management of the pest. And I’ve heard over and over again that if looks as though a chemical in development may cause problems for bees, the company will drop it like a hot potato before it wastes any more money on it.
Practical application: We have only ourselves to blame for the development of resistant pests — it can be avoided by changing the look of our farms, practicing rotations and Integrated Pest Management, developing and planting pest-resistant cultivars, and especially by using pesticides more sparingly.
Farming Practices Continually Change
It’s human nature to be resistant to change, and we have a propensity to rationalize why we can’t do things differently. But farming practices have been changing continually ever since the planting of the first seeds. Hey, a mere hundred years ago, virtually all farming in the U.S. was done with draft animals [[8]]; today farmers are adopting self-driving, GPS-guided, fully automated equipment. Farmers can and will change their pest control methods with time.
For example, the Stockholm Convention in 1991 made of list of the “dirty dozen” persistent organic pollutants — nine of which were pesticides. Since then, the use of such products has been nearly completely phased out in the U.S. [[9]]. California has also reduced the applications of organophosphates and carbamates (which are can be acutely toxic to humans and wildlife, as well as causing neurotoxicological and reproductive issues) (Fig. 4).
Figure 4. Use trend in California of pesticides that are organophosphorus or carbamate cholinesterase inhibitors. This chart shows the reported pounds of active ingredient applied, including both agricultural and nonagricultural applications. Source [[10]].
Not quite there yet: Application of the cholinesterase inhibitors has been reduced, but we are still applying huge amounts of some of them, such as chlorpyrifos, dimethoate, phosmet, carbaryl, and captan. These insecticides have been shown to cause long-term neurological damage to children, as well as being responsible for many serious bee kill “incidents.”
Chlorpyrifos is often the most commonly detected insecticide in bee-collected pollen, and EPA has proposed further restricting it due to the filling of the overall “risk cup” from all the insecticides with the same mode of action. But the ag industry pushed back hard, offering a very good example of how difficult it is to weigh “unreasonable risk to man or the environment” vs. “benefit” to farmers and consumers [[11]], especially when EPA picks out a single product.
Practical application: Pesticides were the most problematic to bees (and the rest of the ecosystem), back in the ‘60s – as much as we hear complaints about pesticides nowadays, in reality the situation is much better (although still far from perfect), and we can expect it to continue to improve. One of the reasons for the shift to neonics is that they appear to exhibit much less neurological risk to humans and other vertebrates.
An example of growers responding to beekeepers
But the writing is clearly on the wall — the ag industry knows that everyone wants to decrease the use of pesticides in general, and especially the most environmentally-harmful ones. Take a look at how the almond industry has responded since beekeepers started reporting dying brood after pollination (Fig. 5).
Figure 5. Active ingredients of insecticides (as opposed to fungicides) applied to almonds. Note how the Almond Board’s issuance of “Best Management Practices” [[12]] has reduced the amount of the insect growth regulators (IGRs) after we discovered in 2013 that they could cause serious brood kills when applied as tank mixes during bloom [[13]]. EPA has now cancelled the conditional registration of flubendiamide, due to its adverse effects in aquatic ecosystems. Thanks to Bob Curtis of the Almond Board of California for the original graph.
Note that the above chart indicates that IGRs were applied to roughly 10% of the one million acres of almonds — often with ugly consequences to the affected hives, which were often not noticed until after the hives had been removed from the orchard. A recent study from Reed Johnson’s lab [[14]] found that Altacor®, when tank mixed with the fungicide Tilt, synergizes to create an even more toxic mix. The Almond Board is getting the word out to their growers to avoid that tank mix.
Practical application: Growers don’t want to kill bees, and if they have a workable alternative, are willing to change their practices. That said, allow me to show you a project that I’ve got going …
Research Project: Effects of fungicides in almonds upon bees
The number one priority of almond growers is to produce a good crop, so if it rains during bloom, they are going to apply fungicides. But since they’re also paying a lot to rent bees to pollinate the crop, and as you saw in the graph above, they’re willing to work with us. What I find lacking is hard data on how the type of, timing of, or adjuvants added to, fungicide applications affect foraging bees. So on a lovely rainy day at the beginning of bloom this season, Huw Evans and I installed entrance flight counters on two hives in each of four different orchards (Fig. 6).
Figure 6. This season I’m making my third attempt to collect flight data from bee counters placed at the entrances of hives in the almonds. We hope to collect hard data on the effect of fungicide applications upon forager behavior and/or loss following a spraying. Photo credit: Huw Evans.
In my two previous attempts to use entrance flight counters, equipment glitches prevented the collection of data. Luckily, technical wizard Huw Evans, who I think of as the Steve Jobs of hive monitoring (due to the user-friendly interfaces that he created for Arnia®), flew over from Italy to install custom-built devices. We owe thanks to Arnia [[15]] and the European research project IoBee [[16]], who helped to sponsor Huw’s trip.
At the time of this writing, the counters appear to be working in good order (Fig. 7).
Figure 7. This pair of hives, fitted with forager in/out counters, scales, a rain gauge, and transmitter are at the U.C. Davis experimental orchard in the “almond city” of Arbuckle (I’ve provided the hives for this research station for over 30 years). Note that the first blossoms are just starting to open. The orchard manager will record the exact times of application of any sprays.
The counters above produce counts of bees exiting and entering over time, as shown below (Fig. 8).
Figure 8. Here’s a snip from the record of one day’s bee flight, with the bee count to the right, and the hour below (we’re also collecting ambient temperature and weight gain data). The black line indicates the number of bees that flew out; the orange line those returning. As expected, not as many return as exit. I hope to see exactly how fungicide spraying affects foraging activity and the return rate.
In 2017, although the flight counters hit a glitch, we were able to collect weight data from a single hive, spanning the period of a fungicide application (Fig. 9).
Fig. 9. Note the strong daily weight gains from the remaining almond bloom on 7 and 8 March, indicating vigorous foraging activity. That foraging appears to have been disrupted by the ground spraying of Bravo fungicide on 9-10 March, suggesting that the bees avoided foraging or were killed, especially on the 10th. Foraging resumed on 11 March.
Although the foraging disruption above occurred after nut set, the weight gain record certainly indicates that the fungicide application had a strong effect upon either bee activity or mortality, and would likely have greatly decreased pollination had it been applied during full bloom. Growers would be interested in such information, and might be willing to adjust their fungicide applications if they felt that it hurt the pollination services that they were paying so dearly for. My hope is that this year’s actual flight data will give us a clearer view of the effect of such sprays, and help us to improve the industry’s Best Management Practices.
Acknowledgements
Thanks to Huw Evans, Pete Borst, Bob Curtis from the Almond Board, and to all the others who have taken the time to answer my questions.
[5] USDA (2011) Research Investments and Market Structure in the Food Processing, Agricultural Input, and Biofuel Industries Worldwide. Economic Research Report Number 130.
[6] Sparks, TC (2013) Insecticide discovery: An evaluation and analysis. Pesticide Biochemistry and Physiology 107: 8–17.
[7] Whitford F, et al (2006) The pesticide marketplace, discovering and developing new products. Purdue University, West Lafayette, IN. (Broken Link!) http://www.ppp.purdue.edu/Pubs/ppp-71.pdf
[14] Wade, A, et al (2019) Combined toxicity of insecticides and fungicides applied to California almond orchards to honey bee larvae and adults. Insects10, 20; doi:10.3390/insects10010020
Allow me to introduce this article with a disclaimer. I do not claim to be a crop farmer nor pesticide expert. But over the past decade, I’ve spent a lot of time listening to those with various points of view — from beekeepers who run their operations on ag lands, conventional growers, and fervent organic farmers, to the ecotoxicologists of the chemical companies, as well as activists and alarmists from all sides of the spectrum. From this overload of information, I’m trying to sort out in my mind how we can aim for win-win solutions. I welcome comments on this continuing deep dive into The Pesticide Situation.
Some Future directions in agriculture
Thomas Malthus long ago pointed out that the human population grows geometrically, whereas our food production tends to increase linearly. But during the last century, three main factors have allowed agricultural production to grow faster than the human population: (1) the invention of the diesel tractor, (2) a declining birth rate, and (3) the “Green Revolution.” During that revolution in the 1950s and ‘60s, new technologies — such as high-yielding crop cultivars, irrigation, and notably, the use of chemical fertilizers and new classes of pesticides ― were brought to agriculture, resulting in marked increases in agricultural productivity. Unfortunately, farmers with large-scale plantings have now embraced those pesticides to the point of dependence, or perhaps even less justified, as a means of risk management.
But the Green Revolution appears to be losing steam, and the specter of mass starvation is looming closer, especially with our warming climate taken into account.
A Decline in Growth in Productivity
Unlike Moore’s Law, in which computers just keep getting faster and faster, advances in agriculture are constrained by both the environment and biology. Somewhat alarmingly, a recent analysis [[1]] detected “sizable and significant slowdowns in the rate of [agricultural] productivity growth in recent decades.” The researchers suggest that the rapid rates of productivity growth enjoyed during the period from 1960 through 1980 were an aberration. This is not good news.
The question then is, will the needs of the human population (as well as its environmental impact) exceed the carrying capacity of the biosphere of our planet within the lifetimes of our grandchildren?
Practical application: We beekeepers, who are already witnessing a shift in some agricultural lands becoming less bee friendly, will be caught in the middle of the struggle for farmers to meet the human demand for their harvests (Fig. 1).
Figure 1. One result of the Green Revolution can be vast landscapes dominated by monocultures — such as by the corn (yellow) and soybeans (green) grown in Iowa farmland above (both being grown mainly as feedstocks, rather than directly for human food). Such monocultures are conducive to the evolution of pest species adapted to utilizing these food sources, as well as surviving any pesticide repeatedly applied to control them. The map for your area can be downloaded at [[2]].
The problem for beekeepers is clearly illustrated in the Northern Great Plains (NGP), on which over a million hives of bees are dependent for summer sustenance — that’s roughly a third of all hives in the U.S. The NGP are undergoing a transition from being a fantastic area in which to keep bees, to being less so. As succinctly stated by Clint Otto [[3]]:
These trends suggest the US beekeeping industry is increasingly reliant on the NGP in providing refuge for honey bee colonies at a time when the carrying capacity of this landscape for supporting colonies is decreasing.
We show a systematic trend in [Conservation Reserve Program lands] loss over the past 10 [years], with the most drastic declines occurring in areas that support the highest numbers of apiaries …
The conversion of CRP to corn and soybeans likely reduces landscape suitability for honey bees because it eliminates important forage lands for pollinators and replaces them with crops of limited forage value. In addition, corn and soybeans are often intensively treated with a variety of agrochemicals that can negatively affect colony health through multiple pathways
Practical application: Beekeepers in the Northern Great Plains are watching the forage for their bees disappear year by year. Thanks to the US Geological Survey for supporting Clint Otto and his collaborators for documenting this, and bringing it to the attention of our legislators.
A Problem with Monocultures
The corn/soy monoculture is a perfect example of how we can create an artificial vegetative landscape that can be maintained only by the intense suppression of competing species. Although we speak of “the balance of nature,” in reality many ecosystems display dynamic swings in populations between the dominant plant species, and the herbivores and parasites that feed upon them. When there’s lots of a specific food plant available, the populations of things that eat that plant may increase until they consume every last one (I’ve witnessed this happen spectacularly). So when farmers plant vast fields of a single plant species, they’re just putting out a huge plate of food for what they term as “pest” insects. In order to maintain that unnatural imbalance, the farmers are then forced to suppress those pests when their populations inevitably respond to that food availability (Fig. 2).
Figure 2. Although expansive plantings of single crops are conducive to high-efficiency farming, such monocultures of a single type of plant create an unlimited supply of food for those species that we then consider as “pests.” The “simple” solution of blasting them with pesticides needs to take into account the unintended consequences upon the rest of us inhabitants of the ecosystem. USDA Photo by: Charles O’Rear.
Conversely, in natural and more diverse ecosystems, it is less likely for any herbivorous insect species to reach huge populations, since that species will tend to be kept in check by its own predators and parasites [[4]]. In a monoculture, that critical diversity of competing insects may be greatly reduced — Schmid [[5]] found only a quarter as many insect species in corn fields as in adjacent prairie land. By providing buffer strips and the rotation of a diversity of crops in an area, “beneficials” may keep pest populations below the economic threshold for damage without the need for pesticides (read a fascinating article on this subject by Jon Lundgren at [[6]]).
Practical application: The removal of livestock and pasture from agricultural ecosystems also eliminates the need for fences. Couple this with larger tractors and Roundup-Ready® plant cultivars, and the landscape loses plant species diversity — thus greatly impacting many other species, including pollinators. From a biological standpoint, diversity in farmland would be strived for, rather than being eliminated.
Below is a different agricultural scenario than the example of Iowa above. Although not yet ideal for Integrated Pest Management, the California farmland illustrated below wouldn’t be as favorable to any single pest (Fig. 3).
Figure 3. Compare this mixed-agriculture setting from California to the previous map — each color represents a different type of crop. There is still pest pressure, but by interspersing and rotating crops, growers can better decrease the rate of pest buildup. On the flip side, such crop diversity may expose bees to a greater array of pesticides as they shift their foraging from one blooming crop to another — this map is less than 7 miles across, so an apiary set in the middle could easily hit every field.
Although it is clear that there are practices that could be adopted by farmers to decrease pest pressure and to ensure the sustainability of production through innovation, the system may be rigged against change due to our current taxpayer-funded Federal Crop Insurance Program [[7]]. The combination of the ethanol mandate coupled with crop insurance, and inadequate returns for land set aside in the Conservation Reserve Program, has in recent years led to the plowing under of millions of acres of land, some of which had formerly been considered as unsuitable for row cropping.
On the other hand, should our ag policies switch from favoring corn for ethanol, to promoting oilseed crops for biofuels (such as canola, sunflower, and soybean), we could perhaps see increased short-term forage opportunities for bees in ag lands, as is occurring in Europe [[8]]. It’ll also be of interest to see whether hemp becomes an important late-season pollen source for bees [[9]]. I’ve checked the amino acid and lipid content of hemp pollen -― it looks pretty good for bees.
Practical application: We can expect agricultural landscapes to continue to change. We beekeepers are now in the position to provide a voice for all pollinators and wildlife in the ag landscape.
The tradeoff in ag areas
Although honey bees are adaptable to a wide range of landscapes — from desert to meadow to deep forest ― the best forage opportunities are typically in “biologically productive” lands, especially at the meadow-forest interface. Due to the fertility of such lands, that’s where much farming takes place.
A recent experiment [[10]] by a group of well-known bee researchers compared colony performance in four carefully-chosen different landscapes in Tennessee ― no ag (forested state park), low ag with urban activity, moderate ag, and high ag (corn, cotton, soy, sorghum, and winter wheat, with the expected exposure to pesticides). I’ve snipped some of their findings below:
It was apparent that bees located in AG [agricultural] areas had access to higher and more sustainable sources of nutrition than those of the [no-ag] area, and starvation losses were only observed in the [no-ag] area. However, mortality of foraging bees resulting from foliar pesticide applications were documented at the [low-ag/urban and moderate-ag] locations on several occasions … [but] these losses did not have measurable impacts at the colony level such as brood production or hive weight …
Pesticide residues in pollen did not occur at levels expected to cause meaningful mortality to honey bees …
[Only] one of 22 pollen samples collected from returning foragers were tested positive for neonicotinoids at trace level …
Exposure to pesticides in colonies of the AG areas did not result in measurable impacts on colony productivity. There may be other effects not measured in this study…
In conclusion, honey bee colonies foraging in moderate and high AG areas were clearly able to grow faster and to a larger size as a result of better access to sustainable nutrition sources than bees foraging in NAG area and a low AG area with urban activity. … Although negative effects of pesticide on colony health were not detected, sublethal doses of insecticides and fungicides were identified in trapped pollen. Better nutrition sources and nectar yields in AG areas helped to develop greater population size, which in turn enabled better colony thermoregulation. [Non-ag] areas may provide a less-toxic environment for honey bees but might not provide sustainable foraging resources, leading to colony starvation. Thus, there appears to be a trade-off between increased food resources and the potential for exposure to pesticides in agricultural systems.
Practical application: The last sentence above says it all — we beekeepers must weigh the tradeoffs. Of course not all agricultural areas are the same — some offer far less “sustainable nutrition.” And in others there are clear impacts from pesticide exposure — I’ve seen heart-wrenching photos of apiaries wiped out by sprays or planting dust, and personally inspected colonies in which the brood was destroyed by a tank mix applied in almonds. Luckily, it’s now hard to imagine that there is any farmer who hasn’t gotten the memo that the public cares about bees.
There are examples of how agricultural practices could be improved in order to reduce pest pressure (as well as numerous other benefits), as exemplified by the farm below (Fig. 4)
Figure 4. One way to slow the evolution of resistant pests is to intersperse various crops and native vegetation, as in this USDA-funded farm on Saipan [[11]]. Each different plant species will harbor different pests and beneficials, which will not only help to prevent the buildup of any one pest, but will create reservoirs of nonresistant individuals of that species — which will then work against the establishment of resistant strains.
Practical application: OK, agronomists and biologists have plenty of suggestions as to how our agricultural practices could be tweaked to be more ecologically sustainable, but what it really boils down to is …
Efficiency, Profitability & Arbitrary rules
Our agricultural practices are continually evolving toward better efficiency and profitability — if a farmer cannot make a profit, he’s unlikely to continue farming (similarly, commercial beekeepers who didn’t adapt to dealing with varroa soon went out of business). But there are also often arbitrary rules to follow — some created by legislators, some self imposed, and some dictated by the market.
For example, a farmer can jump through hoops to become “certified organic,” which prohibits most manmade pesticides or genetically-engineered crops, but allows full mechanization and confers a greater market price for the harvest. On the other hand, Amish farmers (Fig. 5) eschew tractors for cultivation, but may happily use cutting-edge pesticides and genetically-engineered crops. They may not receive more for their harvest, but their costs per acre are so greatly reduced, that they can make a higher net return per acre than with “conventional” farming [[12]].
Figure 5. I took this photo of an Amish farmer working a field as I drove through Pennsylvania. The Amish typically have small farms incorporating a rotation of various crops, livestock, and pasture. This mixed agricultural landscape, although pesticides might be applied, may be more pollinator-friendly than larger monocultures.
What’s becoming clear is that the agricultural community needs to keep its eye on the future. No one wants to cause the extinction of native pollinators, and the ag community needs to maintain a space for beekeepers to fit in. Farmers will adopt some of the latest technology, yet should keep in mind how their forebears farmed before the Green Revolution.
Personal opinion: No one expects U.S. agriculture to go back to the horse and plow, and I fully support “organic” farming [[13]], but as a biologist and beekeeper, I find some of the certification standards for “organic” to be arbitrary, excessive, scientifically unsupported, or even environmentally counterproductive. I suggest that we reward farmers for taking any steps toward improving their practices from an agro-ecological perspective.
The key thing is that farmers should be financially rewarded for making even moderate steps towards ecological sustainability. This can be top down via the Farm Bill, or bottom up from the consumer.
Practical application and action item: The purchasers of all agricultural products respond to consumer demand (hence the rapidly-growing market for “organic”). The average household in the U.S. spends a smaller proportion of its income on food than in any other country in the world [[14]]. Since the payment to the grower is only a fraction of what we pay at the grocery, that means that shifting agricultural practices to be more pollinator-friendly would hardly be noticeable to us at the cash register. Right now is the honey bee’s moment in the sun — the more that we can encourage the consumer to put pressure on the middlemen to reward growers for practicing pollinator-friendly farming, the more quickly things will change.
Next, the problem of pesticide resistance, and some of the coming changes in agriculture that can benefit bees.
Notes and citations
[1] Anderson, M, et al (2018) A century of U.S. farm productivity growth: A surge then a slowdown. American Journal of Agricultural Economics 100(4): 1072–1090.
[2] The USDA Cropland Data Layer interactive map at https://nassgeodata.gmu.edu/CropScape/This informative interactive map shows what’s growing on every acre of the U.S.
[3] Otto, C, et al (2018) Past role and future outlook of the Conservation Reserve Program for supporting honey bees in the Great Plains. PNAS 115(29): 7629-7634.
[4]There are also some cyclical systems, such as those in which “plagues” of locusts periodically graze everything to the ground.
[5] Schmid, RB, et al (2015) Gut bacterial symbiont diversity within beneficial insects linked to reductions in local biodiversity. Annals of the Entomological Society of America 108(6): 993–999.
[8] Breeze, TD, et al (2014) Agricultural policies exacerbate honeybee pollination service supply-demand mismatches across Europe. PLoS One 9(1): e82996.
[10] Alburaki , M, et al (2017) Agricultural landscape and pesticide effects on honey bee (Hymenoptera: Apidae) biological traits. Journal of Economic Entomology 2017: 1–13 doi: 10.1093/jee/tox111
The Federal Insecticide, Fungicide, and Rodenticide Act requires that a pesticide will generally not cause any unreasonable risk to man or the environment — taking into account the economic, social, and environmental costs and benefits of the use of that pesticide. The two italicized caveats are what allow each stakeholder to have a different perspective on pesticide use.
The Pesticide Situation is a contentious topic among beekeepers. There are those who divide everyone else into either of two groups — those who agree with them, or those who are complete morons. I, on the other hand, have found that I learn little in an echo chamber. Indeed, in order to truly understand an issue, one should be able to argue either side’s perspective with equal facility. Thus, I go out of my way to understand the viewpoint of the “other side”.
But rather than simply accepting others’ opinions, I then check out the supporting evidence and scientific interpretation. To that end, in trying to understand The Pesticide Situation, I not only read widely, but also talk to growers, beekeepers, and ecotoxicologists. I thank the many beekeepers who’ve endured my grillings about their pesticide issues, and especially appreciate my conversations with those who are both commercial beekeepers and farmers–who thus see the issue from both sides. The end result is that when I was asked to write an article on pesticides and bees, I soon realized that I couldn’t begin to objectively cover the subject in a single installment—thus you’re reading what’s turned into a series.
So let’s continue by seeing whether I can fairly represent the viewpoints of the various stakeholders—I’m happy to receive emails for suggestions as to things that I may have missed.
The stakeholders
The majority of my readers will be beekeepers; and we of course have a pretty one-sided view about pesticides. But we beekeepers constitute less than a tenth of a percent of the population, and until recent years were generally regarded as weirdos who for some reason kept stinging insects in boxes.
But everything changed when colonies suddenly, and inexplicably, started dying in the early 2000’s. The key word was “inexplicably”, since this caught the media’s (and thus the public’s) attention.
Practical application: although those colony losses were painful to us, it was our good fortune that the sting-ey honey bee was suddenly turned into a beloved poster child for a public coming to terms with the fact that human actions appeared to be possibly threatening that now cute and fuzzy little critter with imminent extinction. Keep in mind that if it weren’t for the demands of the almond industry in California for this non-native invasive insect, our plight might have remained invisible.
The media loved “the sky is falling” story — and it was even better that the death of our bees was apparently due to some mysterious unknown culprit (Fig. 1). Pesticides and the evil chemical companies were obvious suspects, just begging for a lynching.
Figure 1. Fear sells. In preparation for the above issue, the Science Editor of Time phoned and interviewed me for an hour. I carefully explained the reality that honey bees were in no danger of going extinct, and that the number of hives had actually been increasing for a few years. But that didn’t stop the magazine from printing this cover.
Those who demonize the “pesticide companies” should keep in mind that it’s not the companies who actually introduce pesticides into the environment—instead, it is growers, landscape managers, foresters, mosquito control agencies, and homeowners. And those applicators are not that much different from those who keep bees for a living—when we previously pesticide-abhorrent beekeepers were confronted with a pest (varroa) that threatened our livelihood, we suddenly became major pesticide applicators ourselves.
Not only that, but commercial beekeepers in many countries became pesticide scofflaws—desperately applying chemicals to protect their bees from the destructor mite, flouting pesticide regulations, and hoping that they didn’t contaminate their “natural and pure” honey to the extent that the packers would reject it.
My point: Before we go blaming others for being irresponsible with pesticides, we should first (as an industry) look at ourselves in the mirror. From that perspective, perhaps we can better understand the positions of the other stakeholders.
The above said, I’ve found that people of all stripes have a remarkable ability to rationalize whatever it takes for them to perform their job, make a profit, or maintain their style of life. So it’s not so much whether someone is right or wrong, but what the Big Picture effect is of their actions. Unfortunately for the Earth, the effect of 8 billion humans rationalizing their priorities may leave the environment and other species of life on the losing end.
So let’s start with the main applicators of pesticides…
The growers
Rationalization: it’s easy for farmers to rationalize their “need” to apply pesticides, since it’s the “norm” for their peers, and a state agency or salesman may advise them that they need to do it to “protect” their crop. (If you’re not a farmer yourself, you might want to take a look at the sort of pesticide-heavy information that they use for guidance [[1]]).
Hey, farmers gotta pay their bills. No farmer wants to lose his crop to bugs, but must weigh the cost of a pesticide application against what he/she stands to lose. And farmers have choices as to price range and overall ecological toxicity (which may not be clearly explained). Luckily, family farmers live close to the land, and generally want to protect their families and the environment. A face-to-face friendly talk along with a case of honey can really help a beekeeper to get along with his neighbors.
And if we want the growers to be more careful, we gotta show them how to profitably practice pollinator-safe pest management. We can’t just tell them to stop doing what has previously worked for them; we need to give them economically-viable alternatives to the more harmful pesticides. At the governmental and university level, we can help them by providing more demonstration projects and extension outreach to show them how to practice Integrated Pest Management, and to minimize their use of pollinator-unfriendly products.
Politics vs. Science-based Regulation
As with beekeepers, farmers are a minority in this country, and also at the mercy of urbanites. Perhaps the worst thing with pesticides is for politicians (who may know little about agriculture, pesticides, or ecology) to regulate them by fiat, simply in order to appease vociferous urban constituents. Voters can be easily swayed by scare messages, especially by well-meaning but perhaps overzealous environmental groups, or by organic marketers wanting to gain sales by painting others’ products as being dangerous to our health.
Practical application: In the E.U., several countries do not allow the planting of Bt crops, which require fewer pesticide applications. And under public pressure, some have banned some of the neonicotinoid insecticides—summarily forcing farmers to figure out other ways to protect their crops. Many feel that those decisions were based upon politics rather than science, which is hardly the best way to regulate pesticides.
We’re now trying to figure out to what extent the neonic ban cost those farmers, and whether the replacements turn out to be environmentally worse than the seed treatments. As you might expect, the Industry side is claiming massive financial losses [[2]], whereas others are treading more carefully [[3]].
On the other hand, based upon sound science, all regulatory agencies have been trying to phase out the persistent and human-dangerous organophosphates and carbamates [[4]], and California, where I live, may soon ban the most popular one — chlorpyrifos– which is the most frequently found insecticide in bee-collected pollen samples. If it’s indeed banned, farmers will need to adapt.
Speaking of California
California is the nation’s #1 agricultural state, and tends to lead the way in both agricultural practices and pesticide regulation. California voters are also quite health conscious, and leery of any chemical that could possibly be hazardous to their health.
California’s Prop 65
Remember, I’m talking about California, in which every store that sells beekeeping supplies must warn the buyer that woodenware may be contaminated with sawdust, which under Proposition 65 is “a substance known to the State of California to cause cancer.” Ditto for that cup of coffee that you buy on the way home—it’s known by the State to contain acrylamide (which forms any time that foods are roasted). At this point, we Californians are so used to seeing Prop 65 warnings everywhere we go (such as every time we fill up the gas tank), that we just ignore them.
Our Department of Pesticide Regulation (CDPR) maintains a publicly-available database of all applications of restricted pesticides in the State [[5]]. Below is a sample from the 2016 summary, indicating the number of pounds of pesticides applied over time (Fig. 2).
Figure 2. Wow, still a lot of insecticides being applied on California farmland! But keep in mind that the orange fungicide/insecticide plot mostly represents applications of dusting sulfur. Also note that this chart would not show any neonic seed treatments for corn. (There’s not much soy or canola planted in the State). Source [[6]].
CDPR also shows how many acres those pesticides were applied to—for insecticides it was well over a pound per acre of active ingredient. This doesn’t mean that growers can’t successfully shift to safer products. The good news is that over the past 20 years, some of the nastiest “broad-spectrum” insecticides–such as the organochlorines (DDT and chlordane) [[7]], and a number of organophosphates and carbamates have been phased out, whereas inert oil and biopesticide use is increasing (Fig. 3).
Figure 3. The top 5 insecticides applied in California, as far as acres treated [[8]]. Note the trend toward using inert oils (pink). Of interest is the increased use of abamectin (red)—a “natural” and organically-certified insecticide, but still highly toxic to bees (although its residues are short-lived). Also highly toxic to bees are the pyrethroids lambda-cyhalothrin (green line; with a half-life on plants of 5 days) and the neonic imidacloprid (olive line; with an extended half-life on the surface of plants, as well as systemic absorption). Finally, the lepidoptera-specific methoxyfenozide (blue) appears to be pretty safe for bees.
Note that there are 100 million acres in the entire state of California, so the pesticides above are only applied to a tiny percentage of the State acreage. Compare that to the state of Iowa, in which 2/3rds of the state’s total acreage is harvested cropland—virtually all in typically pesticide-heavy corn or soybeans.
How About Human Risk?
With regard to FIFRA, first in the regulators’ minds is the risk of any pesticide to “man”—meaning the consumer, the applicator, as well as those living near agricultural lands. In this matter, California is doing a pretty good job (Fig. 4).
Figure 4. This chart is from Steve Savage’s Applied Mythology website [[9]], which I highly recommend for anyone wanting to get an informed view from a knowledgeable scientist with a practical perspective on sustainable agriculture. Steve and I are both “deeply concerned about the increasingly anti-science environment in which we live today”. Check him out for his entertaining and objectively informative blogs.
Most farmers aren’t organic chemists
California’s legislature passed the Economic Poison Act in 1921, following the inadvertent poisoning of consumers by arsenic insecticides. By 1925, there were about 1,700 products marketed in California for pest control; today there are about 13,000, containing some 1,000 active ingredients. This gives growers a staggeringly-wide choice of chemistries, but unfortunately I doubt that many farmers fully understand each chemical’s ecotoxicological effects. So they often delegate the job to a Pest Control Adviser (PCA).
Pest Control Advisers
Many, if not most, growers depend upon advice by a PCA. Unfortunately, even in pesticide-strict California, one can obtain a license to be a Pest Control Adviser without any educational requirements in biology, chemistry, entomology, or integrated pest management [[10]], so long as they can score 70% in limited testing [[11]]. As pointed out to me recently, PCAs may “just follow the label,” and in almonds, may ignore the published Best Management Practices regarding pesticide applications put out by the Almond Board–thus resulting in unwarranted applications of insect growth regulators (IGRs), pollinator-hazardous “tank mixes,” or the daytime spraying of fungicides onto orchards in full bloom.
A MODERN TALE OF THE FOX GUARDING THE HEN HOUSE: THE INHERENT CONFLICT OF INTEREST THAT EXISTS WHEN PESTICIDE DISTRIBUTORS EMPLOY PEST CONTROL ADVISERS
In California, when agricultural growers want to apply pesticides to their crops, they are required to first obtain a recommendation from a licensed Pest Control Adviser (“PCA”). Imagine you are a grower and in need of such a recommendation. A PCA visits your property to determine what type of chemical you need and how much of it is required to keep your precious crops protected from pests that could destroy your profit. The PCA tells you that he just so happens to sell the exact pesticide that he has recommended for your crop. This is a common scenario experienced by farmers, considering that nearly ninety percent of all PCAs are employed by agricultural chemical distributors and sell the very products they recommend to farmers. …This presents an extraordinary conflict of interest: PCAs employed by pesticide distributors provide pest control advice that is biased toward the profit of their employers and also aimed at earning a commission.
Quoted from the San Joaquin Agricultural Law Review [[12]].
If a PCA is indeed practicing Integrated Pest Management, then they will sample and monitor any pest population to determine whether it is approaching the “economic injury level.” At the point where the pest density exceeds the “economic threshold” (alternatively termed the “action threshold”), control measures are implemented—one option being the application of a pesticide.
Practical application: any PCA is going to err on the side of caution (or perhaps extreme caution) so that they don’t wind up getting blamed for any minor decrease in yield. The net effect is that PCAs who also get commissions from pesticide sales may tend to advise very conservative (and pesticide-heavy) risk management rather than a demonstrated need for treatment (or alternative management strategies). On the other hand, when bee scientist Dr. Gordon Wardell was with Paramount Farms, he advised them that in their environment, there was no need to spray fungicides at all.
Along that line, the need for pollinators does give us one ace in the hole…
Pollination services (one intersection of agriculture and beneficial insects):
Back in 2004, in response to the first major short supply of bees for almond pollination, the offered rental price more than tripled. This resulted in a watershed change for the bee industry—almond pollination was now as important to the industry as was honey production. And although some emphatically blamed the neonics for the elevated colony losses during the CCD epidemic, colony numbers have rebounded, despite a quadrupling of the amount of neonics applied each year [[13]]. As pointed out by a thorough analysis by Ferrier, et al [[14]]:
“High prices are the solution to their own problem.”
Those sky-high payments offered by almond growers for pollination services became the new lifeblood of our industry, with “innovative” beekeepers ramping up their numbers of hives [[15]]. The almond growers now know the value of pollinators—that is, up ‘til petal fall. After that we gotta find somewhere else where our colonies can safely forage. That often involves other agricultural landscapes—and therein lies the rub:
Practical application: we must keep in mind that our major crops, as far as acreage planted—corn and other grains, potatoes, soybeans, and forage crops—aren’t dependent upon insect pollination, so those growers have little reason to care about protecting pollinators on their fields, other than just doing us beekeepers a favor. And if they feel that beekeepers are causing them problems, they’re likely to respond by simply kicking us off their land.
The good news is that there’s a new buzzword in agriculture: “ecosystem services,” which includes not only the pollination performed by native insects, but also the pest control benefits derived from parasites, predatory insects, and birds [[16][17][18]]. This is a good thing for us beekeepers, since what’s good for native pollinators and “beneficials” is also good for honey bees. In order to realize the full benefits of ecosystem services, landowners need to manage diverse habitats at the ecosystem scale, and think twice about any unintended effects from pesticide applications.
Public pressure and Voters
There is always peer pressure. No farmer that I know wants to kill pollinators. And many family farmers are proud to maintain their land in an eco-friendly and sustainable manner, often enjoying having bee hives on their property. Dayer [[19]] points out that “landowners who perceived social acknowledgement of their conservation behaviors were more likely to state an intention to persist after [participating in a conservation] program”(Fig. 5).
Figure 5. Jim and DeAnn Sattelberg received Conservation Reserve Program funding to plant “filter strips” that protect water sources on their Michigan farm. These strips can also provide pollinator and wildlife habitat, so long as they don’t receive too much pesticide drift. Photo credit USDA [[20]].
Unfortunately, at this writing it appears that the 2018 Farm Bill will reduce the Conservation Stewardship Program somewhat, and not provide adequate funding for the Conservation Reserve Program, although some pollinator health provisions may remain in place.
I’m out of space—next month I’ll continue with the future direction of pesticides in agriculture, as well as the perspectives of other stakeholders.
Acknowledgements
Thanks to Peter Borst for library assistance, my wife Stephanie for her patience and proofing, and to my soft-spoken beekeeper/farmer friend Charlie Linder, who delicately sets me straight if he feels that I’ve misinterpreted anything. Feel free to check out discussions at Bee-L.
[3] Kathage , J, et al (2018) The impact of restrictions on neonicotinoid and fipronil insecticides on pest management in maize, oilseed rape and sunflower in eight European Union regions. Pest Manag Sci. 74(1): 88–99.
[4]These neurotoxins are termed acetocholinesterase (ACHE) inhibitors.
[5]Pretty much anything that you can’t get at a nursery, hardware, or garden store.
[7]I was running a farm store when chlordane’s registration was revoked. Although we hadn’t been selling it, our pesticide salesman urged us to stock up. He waxed poetic about how he sprayed his entire property with it each year, and never, ever saw any bugs.
[11]I haven’t attempted to take the test, but the test that I’m required to take every two years in order to maintain my Private Applicator Certificate is not challenging.
[13]As far as I can tell, CCD was the result of a perfect storm of the failure of varroa control products, the evolution of viruses, the invasion of Nosema ceranae, changes in forage opportunities, beekeepers’ failure to manage the mite, coupled with unrealistic expectations for what it takes to supply strong colonies for almond pollination.
[14] Ferrier, RM, et al (2018) Economic effects and responses to changes in honey bee health, ERR-246, U.S. Department of Agriculture, Economic Research Service.
[15]“Innovative” meaning that they stopped pointing the finger, and started controlling varroa and feeding supplemental protein when indicated.
[16] Holzschuh, A, et al (2012) Landscapes with wild bee habitats enhance pollination, fruit set and yield of sweet cherry. Biological Conservation 153: 101–107.
[17] Brittain C, et al (2013) Synergistic effects of non-Apis bees and honey bees for pollination services. Proc R Soc B 280: 20122767
[18] Pfister, S, et al (2018) Dominance of cropland reduces the pollen deposition from bumble bees.Scientific Reports 8: 13873.
[19] Dayer, A, et al (2018) Private landowner conservation behavior following participation in voluntary incentive programs: recommendations to facilitate behavioral persistence. Conservation Letters 11:2 e12394.
I was asked to write an article focusing on pesticides and bees—a subject on which emotions run strong—but are often based upon poorly-informed opinions, one-sided views, or incomplete information. What I hope to do in this series is to help to put things into perspective.
The Earth’s Biosphere
Let’s begin by viewing The Pesticide Situation from the perspective of a “Big Picture” view (Fig. 1).
Figure 1. From this distance, what we see is the Earth’s biosphere—the thin skin on the surface of our planet suitable for life. The biosphere includes the land surface, the seas, and the lower atmosphere. Photo credit NASA [[1]].
The human population
We humans are seriously impacting Earth’s fragile biosphere. The sad fact is that there is absolutely no way that 8 billion humans can live in harmony with nature. The demands of humanity are now exceeding the sustainable carrying capacity of the biosphere–and our population continues to increase at the rate of over 200,000 additional mouths to feed per day [[2]].
We are terraforming biologically-important habitats into agricultural and urban uses, increasing the carbon dioxide content of the atmosphere and the seas, and inadvertently shifting our climate to a warmer (and less life-friendly) temperature. And with regard to this article, we are also chemically polluting parts of the biosphere—which brings us to the subject of bees and pesticides.
Insect populations in general
Although honey bees are not threatened with extinction by any means, they seem to have served as a canary in the coal mine to catch the public’s attention. The reality is that insect populations in general appear to be declining across the globe. But their disappearance doesn’t appear to correlate well with either of the two usual suspects–habitat conversion or pesticides. I’ve spoken with entomologists, and it’s clear that insect species and insect biomass have been declining for some years (even before the first use of any neonics). That said, we certainly still need to pay attention to the effects of pesticides (notably insecticides) upon pollinator populations.
The agricultural situation
Less than three quarters of the land surface of Earth is considered “habitable”—half of which we humans now devote to agriculture of some sort; about 12% to crop production (4.2 million square miles) [[3]]. Max Roser and Hannah Ritchie have nicely illustrated the breakdown in the chart below (Fig. 2).
Figure 2. We use about an eighth of all the habitable land on earth for row and permanent crops. It is upon that portion of the land—plus a small amount for urban and suburban area—that most pesticides are applied. In the case of the U.S, roughly one fifth of all land area is classified as either cropland or urban [[4]]—neither of which serves as good habitat for most insect species. Chart by Max Roser and Hannah Ritchie [[5]].
Worldwide, there are currently about 7 million square miles of land in crop production. Our current human population is nearly 8 billion, which works out to us using about a third of an acre of cropland (on average) to feed and clothe each human being. As we look to the future (Fig. 3), keep that third of an acre per person figure in mind.
Figure 3. In order to meet increased food demand, unless we want to destroy more virgin habitat (thus driving even more species into extinction), we will need to coax more and more calories and protein out of each third of an acre currently used to feed us humans. That means that growers will likely be forced to shift to higher-efficiency farming. Chart credit: farmingfirst.org.
All of agriculture is gearing up for the expected increased demand for food as our human population grows. The ever-expanding middle class calls for more meat, fruit, and vegetables. Unfortunately, the production of those desirable foods requires more acreage per calorie than for the high-efficiency crops: corn (the most efficient), wheat, rice, soy, or potatoes [[6]].
Practical application: luckily, none of those high-efficiency crops are dependent upon bee pollination, and, other than soybeans, are generally not attractive to bees. Unfortunately, the pesticides used upon these crops can still drift onto (or into) plants visited by pollinators.
Much of the Earth’s agricultural land is not being farmed sustainably, and climate change is not helping. Our current reliance on synthetic pesticides is going to change, as pests evolve and we run out of new chemistry options. The big ag companies are well aware of this, and working on more eco-friendly “biologicals,” breeding, and other improvements.
Practical application: pest management is always in a state of change, as pest species develop resistance to each new class of pesticide. Since the registration of new chemistries must now take into account their impact upon pollinators, the future is looking better for bees.
We’re unlikely to go all “organic,” since many farmers may not find the premium paid to be worth the cost [[7]]. But states like California are leading the way, greatly reducing our use of the chemicals of most concern.
The value of pollinators
What I find exciting is that the ag community is starting to realize that the pollinators are an important component of the high-value portion of the agricultural landscape.
Practical application: pollinators encompass a small group of species upon which some of our most favored foods depend—this gives them an economic value, charmingly termed “ecosystem services.” It has not gone unnoticed by activists and fundraisers that this newly-recognized economic value allows us to use the plight of pollinators to gain traction to force the ag industry to start paying more attention to the health of pollinators and the biosphere in general.
We beekeepers caught the media’s attention with CCD, and suddenly the honey bee and the monarch butterfly became the poster children for our need to start protecting pollinators. But we need to ask ourselves…
Who is the real enemy?
The short answer was well put by Pogo creator Walt Kelly: We have met the enemy and he is us (Fig. 4).
Figure 4. Agriculture responds directly to the demands of the American consumer. And we give the farmers a clear directive: we want inexpensive, cosmetically-perfect fruit, vegetables, and meat.Unfortunately, the cheapest way (in the short term) to produce such perfect food requires intensive chemical-heavy agricultural practices. However, we’re learning that many of our current practices are not sustainable for the long term.
We humans are now in a position of needing to decide our place in the biosphere, and how our practices affect the survival of the other species on this planet. As far as our demand for agricultural products, we basically have only three choices:
Our three choices
To greatly reduce the human population. This is a hard sell in the short term.
To convert more natural habitat or grazing land into cropland. At the rate of 200,000 new mouths to feed each day, converting another third of an acre per additional human works out to at least 68,000 acres of habitat conversion per day). This would result (especially in the rainforests) in driving many species into extinction.
To farm existing cropland even more intensively.
Although #3 perhaps sounds distasteful, realistically, it appears to be the best possible solution, and we beekeepers are likely going to have to learn to live with it. The question then is whether that even more intensive farming can allow for the existence of pollinators on that cropland.
Practical application: I’m beginning this series by trying to put things into a realistic perspective (and for us to stop blaming, and instead start being part of the solution). All farmers, large and small, are going to be pressured to intensify production. So as beekeepers, the best that we can do is to accept that fact, and push for progress in figuring out how to allow for pollinators to be an integral part of high-intensity agriculture.
One way to help pollinators in agricultural landscapes is to provide habitat and forage. The other is to minimize the number of them that are killed by pesticides. And this takes us to pesticide regulation, which falls under the purview of…
The Federal Insecticide, Fungicide, and Rodenticide Act
Allow me to briefly summarize the basics of FIFRA [[8][9][10]]:
Generally, before a pesticide may be sold or distributed in the United States, it must be registered (licensed) with the EPA. Before EPA may register a pesticide under FIFRA, the applicant must show, among other things that using the pesticideaccording to specifications, will generally not cause “any unreasonable risk to man or the environment, taking into account the economic, social, and environmental costs and benefits of the use of any pesticide.” I’ve highlighted three critical sections, which give a lot of wiggle room to EPA. And then EPA leaves it up to the States to enforce pesticide use regulations, recordkeeping, and reporting requirements (some states do a better job than others).
The next subject I’ll explore are the perspectives of the stakeholders involved, and what, in their opinions constitute “unreasonable risk,” “environmental costs,” and societal and financial “benefits.”
To be continued…
Acknowledgements
Thanks to Pete Borst for his long assistance in research, and to all the beekeepers, regulators, growers, pesticide applicators, ecotoxicologists, and bee researchers who have taken the time to deeply discuss aspects of The Pesticide Situation with me.
[3] According to FAO definitions, arable land (row crops) accounts for 28.4% of all agricultural land (10.9% of global land area), and permanent crops (e.g. vineyards and orchards) account for 3.1% (1.2% of global land area).
[5] Max Roser and Hannah Ritchie (2018) – “Land Cover”. Published online at OurWorldInData.org. Retrieved from: (Broken Link!) https://ourworldindata.org/land-cover
[6] And also sweet potatoes, leeks, and parsnips. For some interesting reads, check out:
When farmland is scarce, will we all eat roots and tubers?
[7] My opinion on “organic”: I’ve gardened all my life, and resonate with the principle that organic agriculture should be based on the understanding of living ecological systems and cycles, minimal external inputs, improving the soil, and sustainability. In recent years, however, “organic” has become a marketing term for crop and animal production that meets arbitrarily-set restrictions on the use of man-made chemicals or precision-bred crops. Thus, I cannot sell my honey as “organic” since I use paraffin to waterproof my boxes.
Although I have the greatest respect for growers willing to make the effort to obtain “organic” certification, I feel that “organic” has lost its way, and is now simply a marketing term based upon fomenting fear of “chemicals” and “GMOs”—see [[7]]. As a biologist and environmentalist, this bothers me greatly. In order to feed humanity, as well as to protect the biosphere, we need to let go of some of our irrational fear that every chemical is bad (although some clearly are), and positively promote agroecology, rather than just being “anti” this or that.
I suspect that our problems with varroa—at least for commercial beekeepers—may get worse before they get better. But I’d be willing to bet that eventually, we’ll all be keeping bees that are naturally resistant to the mite. It’s quite clear, that left to its own means, Apis mellifera is able to force varroa into the position of being a relatively benign parasite. It is only our current beekeeping practices that are causing this natural evolutionary transition to take so long.
In order to work with our bees to turn varroa into a “benign” parasite (as it is in its natural host Apis cerana), we need to understand what we’ve been doing wrong, and what we can do better. As I’ve tried to explain in previous articles, we’re now dealing not just with varroa, but with the new synergistic coupling of the mite and Deformed Wing Virus (DWV), which I’m calling The Monster. Of interest, it’s only in recent years that parasitologists have started paying much attention to multiparasitism and co-infection [[1]].
When I think of how varroa has turned DWV into the problem that it is today, I love the descriptive terms that Steven Frank [[2]] used to describe competition between genotypes; as you read the snip below, think of how it applies to the varroa/DWV Monster, and the evolutionary forces upon benign or virulent forms of the parasite combination:
There is, however, a problem when two or more genotypes occupy the same host. If one genotype extracts host resources rapidly and reproduces quickly, then the host may die in a short time. A prudent genotype would have relatively low fitness when paired in a host with a rapacious genotype because, for both genotypes, the host is short-lived, and the rapacious genotype reproduces more rapidly than the prudent one [emphasis mine].
Our current beekeeping practices favor the most rapacious genotypes of The Monster by:
Maintaining multi-hive apiaries favorable to the easy dispersal of virus-vectoring mites from collapsing colonies to many other hives, and
Then replacing those fallen colonies each year with more of the same mite-susceptible bee stock.
So long as we continue this hopeless and unsustainable farce, which is completely dependent upon continually-increasing applications of miticides, our dismal rate of 33% annual colony losses is unlikely to improve [[3]] (Fig. 1).
Figure 1. The unfortunately all-to-familiar first signs of a colony entering the death spiral due to the varroa/DWV “Monster.” Note the bee with deformed wings, and the slumping prepupae. Even a strong late-season miticide treatment will not be enough to save this unfortunate colony.
The long-term solution
Most of us are not going to keep smaller apiaries, nor stop replacing deadouts, so the only way that we’re going to solve The Varroa Problem is by focusing upon the genetics of our bees–that is, we need to start demanding that our queen producers get serious about selecting for mite resistance–which would favor the more prudent forms of varroa and DWV.
Practical application: so long as our queen and package producers breed from stock that is dependent upon multiple miticide treatments in order to survive, the situation is unlikely to improve. Only when they start breeding only from colonies that have survived for a year without treatment, will we then stop rewarding the most rapacious forms of the varroa/DWV Monster, and start favoring a more prudent parasite.
The good news is that there are a whole lot of ways that bees can fight The Monster.
Understanding varroa and Apis cerana
The first thing that I’d suggest is for every beekeeper gain an understanding of how varroa’s natural host, the Eastern honey bee (Apis cerana) evolved to keep this parasite in check. It’s only recently, however, that we may have learned one of its critical tactics. It’s always puzzled me exactly why varroa doesn’t attempt reproduce in ceranaworker brood to any extent. Sure, it could be due to the extreme varroa-sensitive hygiene (VSH) of cerana, but that doesn’t fully explain why the mites don’t at least try, or how the nurse bees tell that a pupa is infested.
Three recent studies have greatly helped to clear up the mystery. Lin [[4]] demonstrated that varroa indeed has the ability to successfully reproduce on cerana worker pupae. Page [[5]], showed that the pupae fight back by “social apoptosis”—being entirely intolerant of having mites feed upon them. But the most recent paper was the clincher–Zhang [[6]] discovered that ceranaworker larvae appear to cue on a particular protein in the saliva of the mite, which then triggers the larva to self-sacrifice for the good of the colony. Amazingly, this cue is specific only to late-instar ceranaworker larvae—it has no, or only slight, effect upon any other life stage or sex of either A. cerana or mellifera.
Practical application: as far as I’m concerned, the above is a revelation, and gives us a prime target for selection in our breeding programs [[7]].
With this understanding, I can take a stab at listing the set of rules that Apis cerana sets for the mite:
Apis cerana’s Rules for Coexistence with Varroa
Don’t waste your time trying to reproduce in our worker brood—our larvae will “just say no,” and we adults will try to kill you as we remove our self-sacrificed sisters.
But we’ll offer you a carrot, along with a stick. We’ll allow you to reproduce in our drone brood, under the condition that we’ll maintain a sniffing hole in the capping, and monitor for any signs of stress to that pupa. Hurt it, or transmit harmful viruses, and we will seal you and your bloodline into a waxen grave.
And we’ll further restrict your reproduction by rearing drones only from time to time.
At all times we are going to make your life miserable. We will self-groom and allo-groom (groom our nestmates) with fervor. That means that every time you are forced to shift to a younger worker in order to avoid being carried out by an aging ride that might not return, you’ll have to run the gamut again.
If you play by our rules by being a “prudent” minor parasite, we’ll allow you to vertically transmit to the next generation when we swarm.
By forcing the mites to reproduce only upon the haploid drones (which possess only one allele for each gene), and since only strong drones can catch up with a virgin queen, this confers rather intense selective pressure upon A. cerana genetics, since those colonies that produce a lot of healthy drones have the best chance at getting the genetics of their queen (as opposed to those of the drones that she mated with) into the next generation [[8]] (Fig. 2).
Figure 2. I shot this photo of a drone comet chasing a queen in a commercial mating yard. The queen (apparently no longer a virgin) is at the top left. What appears to be a lucky drone is tumbling upside down slightly below her and to the right.
Practical application: in order to produce and maintain mite-resistant queen lines, our queen breeders will need to shift the genetics of all the drones within flight distance of their mating yards—something that I’m attempting to do over a three-year period [[9]].
The natural evolution of bees towards mite resistance
Apis cerana has figured out a way to live with varroa. Apparently natural selection did not find that it was worth it to eliminate the mite entirely–instead, it relegated the mite to the status of a minor parasite, similar to how we humans allow follicle mites to live on our bodies.
Direction of evolutionary pressure: There is strong natural selective pressure for any wild-living population of Apis mellifera to also work out a deal with varroa—we’ve already seen this happen in South Africa, South America, and in other regional bee populations.
I’d be remiss not to mention varroa tolerance. Honey bees have always been quite tolerant of high varroa levels, so long as DWV or other varroa-vectored viruses are not involved. But some colonies appear to exhibit a high degree of immunity to DWV, even at elevated mite levels. This is an intriguing way for bees to deal with varroa.
Direction of evolutionary pressure: Dr. Eyal Maori has been on the cutting edge of how bees evolve resistance to viruses, and has shown that they can confer resistance via the endogenization of part of the virus genome, as well as via the jelly produced by the nurses [[10]]. We may see some bee strains develop better resistance to DWV.
The future of managed bee populations
But such natural evolution has been inhibited in our commercial stocks–due to our reliance upon miticides, and general lack of hard selection for resistance by most queen producers. On the other hand, a number of breeders who have allowed Bond Selection to take place now observe substantial resistance in their stocks—so long as their bees are kept relatively isolated and away from apiaries of commercial stock. If not kept isolated, the immigration of mites from surrounding apiaries and swarms of the non-resistant bees may overwhelm what would otherwise be an adequate degree of resistance.
This drift problem is exacerbated by the coevolution of varroa and DWV to create the colony-killing Monster about which I’ve written in recent articles. The presence of virulent DWV does not allow for “mite-tolerant” bee stocks, since as soon as their mite level starts to climb, DWV takes the reins and causes the colony to collapse, thus effecting dispersal of the most virulent symbiotic strains of varroa and the virus. I can’t get excited about breeding for “tolerance”—we want to breed for bees that by one means or another, reduce the reproductive success of varroa, at least in the worker brood.
It’s easy to see that the way we’re headed is a dead end street—our dependence upon miticides, and our continual restocking of our apiaries with mite-susceptible stock, favors the evolution of the varroa/DWV Monster. Our current practices are pushing evolution in the wrong direction. Luckily, a number of forward-looking researchers and breeders are working towards solutions, and I feel that the market for queens is about to reach the tip point, where the buyers will start todemand proven resistant stock. At that point, the genetics of our managed bee populations may shift quickly.
Direction of evolutionary pressure: the consumer will likely place strong pressure upon our queen producers in the near future. Once proven mite-resistant stock hits the commercial market, evolution in the bee industry could occur rapidly.
That said, let’s look at which traits we might expect to see evolve…
It’s not necessary to kill a single mite in order to control varroa
Although we beekeepers generally consider the killing of mites to be the most important aspect of varroa management, I suggest that we step back and look at the Big Picture. As evidenced by Apis cerana, it’s not necessary to actually kill a single mite in order to control varroa—the mite’s Achilles Heel is its success at reproduction.
Feel free to play with my varroa model [[11]] and use the “custom” setting to reduce the average number of daughters per foundress per reproductive cycle in half (from 1.45 to 0.72)—at that point, the reproductive success of the mite becomes so low that the varroa population in the hive can’t keep up with its natural rate of attrition (Fig. 3).
Figure 3. I ran four simulations for typical colonies, with the average number of daughters (mated or not) per foundress ranging from 2 to 0.5 (“normal” is around 1.5). Note the differences in the rate of mite buildup. No mites were killed in these simulations—the only holdback upon the buildup of mites was reducing the average fecundity of the foundresses, which can be accomplished by any number of means—several detailed below.
Fortunately for us, it’s becoming clear that the bees have any number of ways for reducing a foundress’s degree of fecundity or reproductive success—I’ll elaborate further on.
Breeding for resistance
Although many beekeepers fancy that they are helping to breed for resistance, simply withholding treatments is a far cry from “selective breeding.” Any meaningful breeding program would require:
A valid assay for actual resistance. This may be difficult in many areas, since colonies exhibiting some degree of resistance may get overwhelmed by mite drift from other hives [[12]].
The breeder needs to control the drone pool, which can only be accomplished by isolation, the flooding of the area with chosen drones, or instrumental insemination.
And then that breeder would need to propagate and disperse thousands of daughters from the resistant queen line.
Realistically, the best that Joe or Jane hobby beekeeper can do is to vote with their dollars—by supporting those breeders who are indeed making a serious effort at selecting for resistance.
Practical application: it’s hard for me to get excited about breeding for mite tolerance—simply because colonies with a heavy mite load will always be more stressed. So I feel that we should focus upon selecting for resistance to mite buildup. We don’t initially need completely mite-proof bees; partially-resistant colonies that required only a single treatment a year would be a big step in the right direction.
That said, I’ve bred bees for certain traits for many years, and generally found it pretty easy to breed for color, temperament, productivity, and resistance to AFB and tracheal mite. I was completely surprised by how difficult it’s been to breed for bees that can handle varroa. I figured that if Apis mellifera came from an Asian heritage, as has long been assumed, that it would still possess deep in its genome some tools for fighting mites—but if that were the case, it shouldn’t be so hard to tease those resistance traits back to the forefront. I mean, what gives—it took only about five years for bees to develop widespread resistance to the tracheal mite.
A possible explanation for this conundrum has been recently suggested by Dr. Keith Delaplane, based upon a revision of the course of migration and evolution of the genus Apis by Kotthoff [[13]]. It may be that our Western honey bee actually split off as a sideline of the genus before the other honey bee species evolved in Asia—where they were only then exposed to parasitic mites. Thus, Apis mellifera may have never needed to assemble a toolbox for fighting varroa, and was blindsided by the mite when it later jumped host from the bees’ mite-resistant cousin.
But that certainly doesn’t mean that our bees can’t repurpose their existing tools to do so. There are any number of possible ways that the honey bee can fight back against varroa—the ones that we’ve already identified are well reviewed at [[14]]. Here’s a quick rundown on some promising traits to select for:
Grooming and biting
The first mite-resistance trait that often comes to mind is grooming/biting behavior. And although it makes us feel good to imagine our bees crushing mites in their jaws, I find little compelling evidence that they actually kill healthy adult mites very often, and suspect that most mites that get crushed were already too feeble to get away [[15]]. So although we clearly want bees that groom mites vigorously, I doubt that grooming alone will be enough to solve The Varroa Problem [[16]]. So selection for grooming, yes–but along with selection for other more effective traits.
Entombment under the cocoon
Update: Dr. Jeff Harris, working at the Baton Rouge lab, found that in one mite-resistant bee stock, that the 5th-instar larvae were able to kill foundress mites by trapping them under the silk as the larva spun its cocoon. Note: this would be a very easy trait to assay for, by simply inspecting the cocoons in the cells of any colonies exhibiting low mite counts. See Jeff’s video, in which he mentions this trait, at https://www.youtube.com/watch?v=a2vg59Snt6c
Postcapping duration
Varroa enjoys far greater fecundity (mature daughters per foundress) in drone brood than in worker brood, due to the longer postcapping duration for the drone pupae (15 rather than 12 days) (Fig. 4).
Figure 4. The longer postcapping duration of immature drones allows for more daughter mites to successfully complete their development. This allows for a theoretical average emergence of roughly 4 daughters emerging from a drone cell vs. 1.45 daughters per worker cell. The actual number of mated daughters per foundress, however, may be considerably less, especially when more than one foundress invades a cell. Image courtesy Dr. Stephen Martin [[17]].
Some early research by Büchler [[18]] suggested that selecting for shorter postcapping duration might be constructive, and a number of teams have since tried this approach, but generally not found it to be of substantial benefit. However, a recent study by Oddie [[19]] found what appeared to be a slight benefit, which in conjunction with others resistance traits, might help to do the trick.
Biological observation: bee immatures develop more quickly if broodnest temperature is raised a bit. It’s not clear whether varroa responds similarly, since the mite appears to reproduce better at lower temperatures. I see no reason that Apis mellifera couldn’t use broodnest temperature regulation to its advantage.
Varroa-sensitive hygiene
Colonies may differ greatly in the degree of diseased brood hygiene that they exhibit, which is surprising, since rapid hygienic removal of sick brood is a very effective way for colonies to control diseases [[20]]. It’s very easy to select for strong hygienic behavior by using the freeze- or pin-killed brood test. This trait will also confer some degree of mite resistance, since mite- or virus-sickened, or abnormally-developing brood may emit odors that initiate generic hygienic removal (Fig. 5).
Figure 5. These bees have uncapped, and are chewing out apparently-abnormal pupae. This lack of tolerance for abnormal development or olfactory signals is a well-proven means for bees to prevent the mite from reproducing.
Even more specific is some sort of varroa-sensitive hygiene (VSH). Apis cerana exhibits VSH to the extreme, due to the self-sacrifice of its worker larvae. VSH is a generic term for any sort of hygienic behavior that by some means specifically targets mite-infested brood cells or cells in which mites are reproducing (generally specific to the worker cells), and is a well-proven trait–a colony that exhibits even 50% VSH can pretty much keep varroa in check (Fig. 6). The USDA VSH and Pol lines, as well as the Russian bees all exhibit strong VSH.
Figure 6. The strong effect of VSH upon varroa reproductive success. (A) A simulation for mite buildup in a colony not exhibiting VSH; (B) The result of a 50% rate of VSH. I suspect that my model currently underestimates the full effect of VSH, since it does not yet account for an increase in mortality of the disturbed foundress.
Practical application: VSH is a well-proven and effective trait for mite resistance.
I hesitate to mention specific queen producer names, but open-mated Minnesota Hygienic queens are commercially available, and a number of California breeders have also been selecting for freeze-kill hygiene. Certain breeders offer purebred Primorsky Russian stock (outbred stock may not perform the same). Dr. Albert Robertson from Canada is collaborating with U.S. producers to make his Saskatraz line available in the US. Other breeders may also offer bee stock selected for resistance, but as yet there is no testing organization to verify their degree of resistance.
Practical application: our industry needs to start an “All-America” type of organization of volunteer testers to evaluate stocks with claimed mite resistance.
An exciting development is that the USDA-ARS, Project Apis m, and Arista Bee Research are collaborating with a large breeder to make VSH production queens available on large scale—perhaps by next year. This stock will indeed have hard mite-resistance data to back it up.
Along with VSH there is also the uncapping/recapping trait, which may include the creation of “bald brood” (Fig. 7)—we are still working on how to better understand this trait [[21]].
Figure 7. Many of us have observed “bald brood” in our colonies—something that we didn’t notice often prior to varroa. Perhaps such uncapping (apparently sometimes only temporary) may disrupt mite reproduction, or desiccate the unsclerotized male or immature female mites.
Other traits that may confer resistance
VSH is effective, but it’s also pretty ugly and wasteful, since it involves the death and physical removal of the brood involved. Something that I find fascinating is what the ARS used to call SMR—Suppression of Mite Reproduction. SMR appears to be the main resistance mechanism of the African savannah bee (A. m. scutellata), for which beekeepers find no need to control varroa. As straightforwardly noted by Nganso [[22]]:
…hygienic and grooming behaviors did not significantly differ between subspecies [African and European] with respect to Varroa mite-infestation levels recorded, suggesting that other resistant mechanisms such as suppression of mite reproductive success and/or lower viral prevalence within honeybees and mites might play an important role in honeybee responses to mite infestation.
This is also something that appears to have been selected for by some populations of resistant bees in Europe (well reviewed by Panziera [[23]]. We’re watching evolution in action—and our bees, given the chance, may surprise us by coming up with different ways to deal with the mite. Unfortunately, intentional selection for SMR requires tedious examination of the brood (Fig. 8).
Figure 8. I’m removing mature brood one at a time, under a dissecting ‘scope (the spotlight is turned off for the photo). Such dissection can be used to determine whether there was a mite in the cell, and whether it reproduced. Yes, tedious.
Kairomones and proteins—Achille’s heels of varroa
Other potential resistance mechanisms have to do with olfactory cues used as kairomones by varroa. A foundress mite must start (and then end) two critical, yet separate reproductive processes, carefully cued and timed to coordinate with the timing of the development of its immature bee host. The first step is oogenesis (the creation of the egg cell), followed by vitellogenesis (yolk formation, which is also dependent upon the mite’s feeding on the larva’s fat bodies). Oogenesis is triggered by one or more olfactory cues from the late-instar bee larva [[24][25]]. And somehow, the foundress is cued to stop producing eggs five days after the larva pupates, despite the fact that the pupa still has four days before it will emerge.
These cues are absolute requisites for successful mite reproduction, plus, the cues must cause the mite’s first egg to be a haploid male (which in varroa, surprisingly comes from a fertilized egg) [[26]]. The tweaking of any of these cues by the bees has the potential to mess up successful reproduction by the mite.
Now here’s where it gets interesting. Although varroa has a larger genome than does the bee, it appears to have given up the ability to produce some critical proteins—instead assimilating them directly from its bee host without digestion. Although discovered by Tewarson over 25 years ago [[27]], only recently have molecular biologists started looking into this fascinating aspect of varroa biology, publishing a protein atlas for varroa by McAffee [[28]] and an informative breakdown by Mondet [[29]], by which we may figure out some of the mite’s weak spots.
One recently-found possibility is suggested by Conlon [[30]], who notes that mite vitellogenesis is triggered by the molting hormone ecdysterone (ecdysis is the scientific term for molting). He notes that:
[Varroa’s reproductive] pathway is incomplete with only three of the seven genes from the ecdysone biosynthetic pathway present in the V. destructor genome… Functional forms of ecdysone are capable of ingestion by Varroa; suggesting the reduced number of genes may be an adaptation of the mite to its parasitic lifestyle and missing compounds are acquired through its haemolymph diet. This raises the possibility that the pulse of prepupal ecdysteroids is not a signal but a necessary physiological component for the successful initiation of reproduction in V. destructor.
The reason that Conlon’s suggestion is of great interest is that his research suggests that two independent populations of mite-resistant bees may both be downregulating their ecdysone-linked genes, thus possibly conferring resistance at the molecular level of the mite’s reproductive pathway [[31]].
Evolution is not limited by our imagination. And this is the reason that I’m curious to see what happens if I simply apply strong selective pressure upon the breeding population of my own operation, based upon alcohol wash count monitoring alone (plus them mothering productive, gentle colonies). Who knows what traits they might come up with?
Acknowledgements
Thanks to Peter Borst for his assistance in literature search, to all the bee researchers I’ve spoken with on this subject, and to my wife Stephanie for her suggestions on my manuscript.
References
[1]Furthermore, the reality, more or less ignored until recently, is that most parasites co-occur with other parasites. Vaumourin, E, et al (2015) The importance of multiparasitism: examining the consequences of co-infections for human and animal health. Parasites & Vectors 8:545 Open access.
[2] Steven A. Frank, SA (1996) Models of parasite virulence.The Quarterly Review of Biology 71(1): 37-78.
[4] Lin Z, et al. (2018) Reproduction of parasitic mites Varroa destructor in original and new honeybee hosts. Ecol Evol. 2018:1–11.
[5] Page, P, et al (2016) Social apoptosis in honey bee superorganisms. Nature Scientific Reports 6:27210
[6] Zhang, Y & R Han (2018) A saliva protein of varroa mites contributes to the toxicity toward Apis cerana and the DWV elevation in A. mellifera. Scientific Reports 8:3387.
[7] It would be a relatively simple bioassay or gene for Marker-Assisted Selection.
[8] Since, other than some recombination, the haploid drones carry only the genetics of their mother.
[9]It’s easy to talk the talk; I’m walking the walk, and openly sharing my methods, successes, and failures.
A spoiler—we’re pretty excited about what we’re seeing so far in our November mite washes of our “potential breeders” this season.
[10] Maori, E., et al (2007) Reciprocal sequence exchange between nonretro viruses and hosts leading to the appearance of new host phenotypes. Virology 362 (2): 342-349.
Garbian Y, et al (2012) Bidirectional transfer of RNAi between honey bee and Varroa destructor: Varroa gene silencing reduces varroa population. PLoS Pathog 8(12): e1003035.
[12] This was a problem during ARS’s development of the Primorsky Russian stock.
[13] Kotthoff U, et al (2013) Greater past disparity and diversity hints at ancient migrations of European honey bee lineages into Africa and Asia. Journal of Biogeography. 40(10):1832-1838.
[15] My varroa model indicates that if the bees were able to increase the baseline varroa mortality rate from 0.5% a day to only 2% a day, that varroa would barely be able to survive in the hive. Since mites can be easily found in colonies selected for “biting,” I must question whether this trait has substantial effect.
[17] Published in Munn & Jones, ed (1997) Varroa! Fight the mite. Conference proceedings, International Bee Research Association.
[18] Büchler, R & W. Drescher (1990) Variance and heritability of the capped developmental stage in European Apis mellifera L. and Its correlation with increased Varroa jacobsoni Oud. infestation, Journal of Apicultural Research, 29(3):172-176.
[19] Oddie, MAY, et al (2018) Reduced postcapping period in honey bees surviving Varroa destructor by means of natural selection. Insects doi:10.3390/insects9040149
[20]The question is, if rapid hygienic behavior is of general benefit to the colony, we would expect evolutionary pressure to have selected for all bees to be “hygienic.” Why that’s not so, I don’t know. See:
Al Toufailia, H, et al (2018) Both hygienic and non-hygienic honeybee, Apis mellifera, colonies remove dead and diseased larvae from open brood cells. 373: 1751.
[21] Oddie, M, et al (2018) Rapid parallel evolution overcomes global honey bee parasite. Scientific Reports 8: 7704.
[22] Nganso BT, et al (2017) Hygienic and grooming behaviors in African and European honeybees—New damage categories in Varroa destructor. PLoS ONE 12(6): e0179329.
[23] Panziera, D, et al (2017) Varroa sensitive hygiene contributes to naturally selected varroa resistance in honey bees. Journal of Apicultural Research 56( 5): 635-642. Open access.
[24] Garrido, C & P Rosenkranz (2004) Volatiles of the honey bee larva initiate oogenesis in the parasitic mite Varroa destructor. Chemoecology 14: 193–197.
[25] Frey, E, et al (2013) Activation and interruption of the reproduction of Varroa destructor is triggered by host signals (Apis mellifera). Journal of Invertebrate Pathology 113: 56–62.
[26] Sabelis, MW & CJ Nagelkerke (1988) Evolution of pseudo-arrhenotoky. Experimental & Applied Acarology 4: 301-318.
[27] Tewarson, NC & W Engels (1982) Undigested uptake of nonhost proteins by Varroa jacobsoni, Journal of Apicultural Research 21(4): 222-225. Since unaltered bee proteins are directly utilized by varroa, this is a potential way for bees–or beekeepers–to sneak something disruptive into the mite, such as RNAi or some other attached chemical.
[28] McAfee, A, et al (2017) A Varroa destructor protein atlas reveals molecular underpinnings of developmental transitions and sexual differentiation. Mol Cell Proteomics 16(12): 2125–2137.
[29] Mondet, F, et al (2018) Transcriptome profiling of the honeybee parasite Varroa destructor provides new biological insights into the mite adult life cycle. BMC Genomics 19:328.
[30] Conlon, BH, et al (2018) A modified honey bee Ecdysone pathway inhibits reproduction in Varroa. In Of Mites and Men: The independent evolution of host-induced Varroa infertility in the drone brood of Apis mellifera. PhD Dissertation, Martin-Luther-Universität.
[31] Some astute research groups will likely apply to patent varroa-control products based upon suppressing the production of mite-required proteins by the bees, or by using those undigested proteins as transporters of a bound chemical.
The Greek philosopher Heraclitus maintained that there is nothing permanent except change. This certainly applies to biology and the business of beekeeping, both of which are always in some state of evolution. However, beekeeping in the U.S. was unusual in that it changed little between the mid 1800’s and the arrival of a slew of new parasites starting in the late 1900’s. Since then, it’s been hard to keep up with the changes. As a biologist, it’s been fascinating to watch evolution before my eyes; as a beekeeper, it’s been, shall we say, “challenging.”
Caught up in the immediacy of our everyday lives, we humans often have a difficult time with grasping the long-term consequences of our actions. This has certainly been the case since the invasion of varroa, which we’ve dealt with by one stopgap measure after another. It’s frustrating to admit that after three decades, the mite continues to remain our most pressing problem.
It’s important to keep in mind that we’re in the middle of major evolutionary changes in the business of beekeeping, as well as the rapid biological coevolution of the honey bee, the varroa mite, and its symbiotic partner–Deformed Wing Virus (DWV). There’s no way to predict the future, but I’d like to wrap up this series on The Varroa Problem by tying up some loose ends, and pointing out some likely directions of the ongoing evolution of the players.
The Evolution of varroa
The varroa mite may be only the size of a pinhead, but the tiny parasite possesses a genome twice the size of that of the honey bee [[1][2]]. And it has demonstrated its ability to rapidly evolve and disperse traits that confer it increased fitness—remember how it took only about three years for most of the U.S. population of varroa to gain resistance to coumaphos?
Practical application: varroa has clearly demonstrated its ability to rapidly evolve, and our current beekeeping practices ensure that any successful mutants will get quickly distributed.
The fact is that varroa is well-adapted to parasitizing the drone pupae of its natural host Apis cerana, but is still under selective pressure to better adapt to its recent host shift—in which it primarily reproduces on the worker pupae [[3]].
Direction of evolutionary pressure: we can expect this parasite to continue to evolve its behavioral and physiological responses to cues from worker brood of Apis mellifera, adapt to the higher temperature and shorter postcapping period of that worker brood, as well as to develop further resistance to miticides. We can likely expect varroa to become even more “virulent” than it is now.
And varroa has only begun to exploit its symbiotic coevolution with DWV—a marriage that has created a “Monster” that now acts more like a parasitoid than a parasite, killing its host colony towards the end of the season in order to benefit from greater dispersal to other hives. Rather than it being to the mite’s advantage to be a relatively benign, vertically-transmitted parasite–as it is in its natural host–it may now be to the mite’s advantage for each female to complete a reproductive cycle or two, and then to try to hitch a ride to another hive [[4]].
Update and Clarification: I need to make clear that I’m suggesting that we must consider what we define as the host or hosts of DWV, with regard to the honey bee. The virus can also infect other insect species, but evolution will select for strains of the virus best adapted to those species—thus I’m going to ignore those host species. The hosts relevant to this discussion are honey bee larvae, bee pupa, bee adults, and the honey bee colony as a whole. Some researchers claim that DWV also appears to reproduce in the body tissue of the varroa mite, but all but one of their studies were based upon the circumstantial evidence of detecting either positive strand DWV (indicating presence) or negative strand DWV (indicating reproduction) in whole-mite extracts, which typically include virus-laden honey bee fat body tissue. On the other hand, a number of studies [] strongly suggest that DWV does not appear to replicate in actual varroa body tissue [[5]]. The only study that I’ve seen to date (February 2019) that appeared to find DWV actually replicating in mite tissue was by Campbell [[6]], and then only in mite brain tissue – unlike the virus’ replication in several tissue types as in its other hosts, so I am still unconvinced that varroa is anything but a vector of DWV. This is an important consideration, since if varroa does not suffer from being infected by DWV, then there is no direct cost to it from vectoring it, and perhaps a major benefit.
This brings us back to the four remaining honey bee hosts of DWV – the larvae, pupae, adults, and the colony as a whole. Recent research [[7]] indicates that DWV, whether acquired by feeding or injection, is relatively benign to bee larvae and pupae, and numerous studies indicate that adult bees carry a covert infection by the virus [[8]]. Thus, at the colony level, DWV virus appears to be a relatively benign parasite.
But when varroa enters the picture as a vector, things change. Adult bees and brood start to die — although it’s not yet clear to me what is actually causing the brood to die. But more important, the entire host colony then dies, but before then, drifting bees horizontally transmit DWV-vectoring mites to other host colonies in the vicinity. If they didn’t do so, there would be a huge fitness cost to both the mite and the virus from killing the host colony. Since the honey bee as a species (as well as we beekeepers) are more concerned about the death of entire colonies than of individual bees, we need to focus upon how the mite-virus parasitoid is evolving to cause the death of its host at the colony level. The take-home message is that this would not be favored by natural selection unless humans kept artificially replacing the fallen host colonies.
Evolutionary note: in A. cerana, it is the genes of mites that transmit vertically via swarms from healthy colonies that get into the next generation of host colonies. Currently in A. mellifera, few mite bloodlines are transmitted vertically via purchased queens or packages—rather most genetic transmission is via bee drift from, or the robbing of, collapsing colonies. Take home message: beegenetics may be largely controlled by the commercial producers, but mitegenetics evolve in response to our beekeeping practices.
The take-home from this is that the current evolutionary strategy of varroa appears to be:
Start in the spring with an assortment of near-clonal matrilines of the mite in each hive,
For those inbred strains to race against each other to produce the most offspring during colony buildup in the spring and summer, and then,
To then cross-mate within the hive with other successful strains [[9]] just before the mites, in conjunction with DWV, kill their host colony, and then,
For the most numerically-successful mite/DWV strains to catch rides on exiting bees to other hives in the neighborhood.
By this strategy, the most successful mutations can quickly shift the genetic structure of the local mite/virus population, and we humans, with our motorized transport of the mite all over the continent, ensure that those strains get quickly and widely dispersed.
Direction of evolutionary pressure: We are currently on track to witness varroa continuing to coevolve with DWV to become an ever more effective colony-killing parasitoid.
The evolution of DWV
DWV has long been around as an insect virus, but was rarely noticed as being a problem to honey bees until it hooked up with varroa [[10]] (Fig. 1). The two species of parasite have now “figured out” how to work closely together in a mutualistic manner [[11][12]], and can be viewed as a single threat [[13]].
Figure 1. We normally observe the ugly signs of DWV only if a colony is highly infested with mites. But that doesn’t mean that the virus isn’t there—most all bees in a hive nowadays carry DWV. It’s the combination of varroa and DWV that is deadly.
As occurred with the invasion of Nosema ceranae replacing N. apis, a new strain of DWV (Type B) appears to be replacing the existing predominant Type A strain of the virus in this country [[14]], but it’s too early to tell what the effect will be.
Research needed: of great interest to me is that, although varroa and DWV are now closely linked, I’ve yet to see compelling evidence that the virus actually infects and reproduces in the mite’s body tissues (but there is strong suggestive evidence otherwise). This question of whether varroa acts solely as a vector, or rather as an amplifying intermediate host, begs for further clarification.
DWV exists as a continually-mutating “cloud” of variants, with several strains often being found within a single hive or apiary. A single strain may dominate for years in an area, or be replaced by another [[15]]. It’s not yet clear as to whether coinfection with more than one strain of the virus is more detrimental to the bee. The thing to keep in mind that the virus has the capacity to evolve far more rapidly than either the mite or the bee, due to its very high mutation rate and quick generation time (hours rather than weeks or years).
I was recently speaking with Dick Rogers of Bayer about his observations that in some hives the workers may take more than 12 days to emerge after being capped over. This brought to mind something that (now-doctor) Judy Wu pointed out some years ago [[16]]—that bees reared in miticide/pesticide-contaminated combs may exhibit a longer post capping period, thus possibly resulting in greater fecundity for varroa, since an extra day might allow the maturation of an additional daughter.
Direction of evolutionary pressure: it’s not yet clear to what degree our miticide residues and other chemical contamination of the combs play a part in this, but it does point out a potential direction for DWV evolution—if DWV could manage to delay the development of worker pupae by a day or two, it would nearly double the reproductive success of the mite. Scary, huh?
And that doesn’t even take into account how DWV (and some other viruses) apparently benefit from the suppression of the bee antiviral immune response by at least some insecticides [[17]]. Let me make clear though, that in my own operation, with zero exposure to neonics, DWV is still the main problem if I don’t keep varroa under check.
So long as beekeepers fail to control the level of mites in their hives (Fig. 2), we can expect DWV to be a major player in colony health and survivorship.
Figure 2. I overlaid estimated alcohol wash counts (and some arrows) over the results of the USDA National Honey Bee Disease Survey results [[18]]. Note how poor a job that beekeepers in general are doing at keeping varroa levels below the orange economic harm line—above which DWV typically starts to become problematic.
Direction of evolutionary pressure: DWV has quickly adapted to take advantage of its mutualistic relationship with varroa. The virus has now “learned” how to use varroa as a vector to spread itself to other hives. So far, common beekeeping management practices have been playing into the virus’ hands. I hesitate to guess the future, but wouldn’t bet against DWV becoming even more problematic.
The evolution of our beekeeping practices
Over the past three decades, in my own operation, I’ve gone from a hands-off approach of just letting the bees do their own thing, to now alcohol washing every single hive in the operation, and applying organic acids or thymol at least four times a year (but I’ve also gone from getting $12 for almond pollination to over $200). Prior to varroa hooking up with DWV so effectively, it was no problem to keep 72 hives in a yard. Nowadays, that would be foolish unless one really stays on top of the mite.
The reason is the well-documented drifting of bees between hives (Fig. 3), and if those bees are carrying mites and DWV, that’s a problem. Evolutionarily, late-season drift of mite-infested bees as colonies collapse from DWV widely disperses the most virulent strains.
Figure 3. How many mite-carrying drifted bees are entering your hives? At this hive entrance, the guards are checking out a drifted bee—which will likely be eventually accepted into the hive. Even if not, there’s little to prevent a mite from jumping onto one of the guards during the scuffle.
A sneak peek: I’m currently running a large field experiment in which we’ve tagged over 5000 bees in varroa-infested hives, and tracked where those bees wind up. I can already share that there is plenty of drift to hives over 500 feet away, and a fair amount of drift to an apiary a half mile distant.
Direction of evolutionary pressure: so long as beekeepers run non mite-resistant stock and have colonies crashing from DWV in late summer and fall, we will continue to reward the Monster for killing our hives, so long as we keep replacing those unfortunate colonies with yet another meal of the same sort of non-resistant bees.
Evolution of treatments
The writing was on the wall from the beginning—mites have an amazing ability to evolve resistance to synthetic miticides. We’ve gone through several already, and much of our industry is now hanging by the thin thread of amitraz. In country after country, beekeepers are being forced to learn how to use organic acids and thymol instead. We can cross our fingers that some miracle in pheromones, botanicals, biocontrols, olfactory blockers, or RNAi is going to save us, but I know of nothing very promising in the pipeline.
Direction of evolutionary pressure: switching to integrated pest management of varroa, using “natural” treatments, will certainly buy us time, but there is no question that the ultimate solution to The Varroa Problem will be a wholesale shift to running bee stocks that are innately resistant to the mite.
Practical application: my sons and I have run a successful small commercial operation since 2001 without using any synthetic miticides. It can be done. If your operation is completely dependent upon amitraz, I suggest that you start by practicing with a Plan B before you are forced to do so. And start demanding that your queen producers get serious about selecting for mite resistance!
To be continued…
Acknowledgements
Thanks to Peter Borst for his assistance in literature search, to all the bee researchers I’ve spoken with on this subject, and to my wife Stephanie for her suggestions on my manuscript.
References
[1] Cornman, R. S. et al. (2010) Genomic survey of the ectoparasitic mite Varroa destructor, a major pest of the honey bee Apis mellifera. BMC Genomics. 11: 602.
[2] The Honeybee Genome Sequencing Consortium (2006) Insights into social insects from the genome of the honeybee Apis mellifera. Nature. 443: 931-949.
[4] Nolan, MP IV (2016) Impacts of inter-colony distance, mite host choice, and colony polyandry on the host/parasite relationship between Apis mellifera and Varroa destructor. Dissertation, University of Georgia.
[5] Ongus, JR (2006) Varroa destructor virus 1: A new picorna-like virus in Varroa mites as well as honey bees. Thesis, van Wageningen Universiteit.
Todd JH, et al (2007) Incidence and molecular characterization of viruses found in dying New Zealand honey bee (Apis mellifera) colonies infested with Varroa destructor. Apidologie 38:354–367.
Fievet, J, et al (2006) Localization of deformed wing virus infection in queen and drone Apis mellifera L. Journal of Virology 3: 16.
Zhang, QS, et al (2007) Detection and localisation of picorna-like virus particles in tissues of Varroa destructor, an ectoparasite of the honey bee, Apis mellifera. J. Invertebr. Pathol. 96: 97–105.
Santillan-Galicia, MT, et al (2008) Immunolocalization of deformed wing virus particles within the mite Varroa destructor. Journal of General Virology 89: 1685–1689.
Ryabov, EV, et al (2014) A virulent strain of deformed wing virus (DWV) of honeybees (Apis mellifera) prevails after Varroa destructor mediated, or in vitro, transmission. PLoS Pathogens 10, e1004230; doi: 10.1371/journal.ppat.1004230.
Erban, T, et al (2015) In-depth proteomic analysis of Varroa destructor: Detection of DWV-complex, ABPV, VdMLV and honeybee proteins in the mite. Scientific Reports 5:13907.
Nordstrom, S. (2003). Distribution of deformed wing virus within honey bee (Apis mellifera) brood cells infested with the ectoparasitic mite Varroa destructor. Exp Appl Acarol 29, 293–302.
[6] Campbell, EM, et al (2016) Transcriptome analysis of the synganglion from the honey bee mite, Varroa destructor and RNAi knockdown of neural peptide targets. Insect Biochemistry and Molecular Biology doi: 10.1016/j.ibmb.2015.12.007.
[7] Tehel, A, et al (2019) The two prevalent genotypes of an emerging infectious disease, Deformed Wing Virus, cause equally low pupal mortality and equally high wing deformities in host honey bees. Viruses 11: 114.
[8]Reviewed by de Miranda JR, Genersch E (2010) Deformed wing virus. J Invertebr Pathol 103:S48–S61
[9] Beaurepaire, AL, et al (2017) Seasonal cycle of inbreeding and recombination of the parasitic mite Varroa destructor in honeybee colonies and its implications for the selection of acaricide resistance. Infection, Genetics and Evolution 50: 49–54.
[10] Martin, SJ., et al (2012) Global honey bee viral landscape altered by a parasitic mite. Science 336:1304-6.
[11] Di Prisco, G, et al (2016) A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health. PNAS vol. 113(12): 3203–3208.
[12] Erban, T, et al (2015) In-depth proteomic analysis of Varroa destructor: Detection of DWV-complex, ABPV, VdMLV and honeybee proteins in the mite. Sci Rep. 5: 13907.
[13] For some reason, DWV doesn’t appear to be much of a problem in mite-infested colonies in Africa.
[14] Ryabov, EV, et al (2017) Recent spread of Varroa Destructor Virus-1, a honey bee pathogen, in the United States. Scientific Reports 7: 17447.
[15] Jamnikar-Ciglenecki, U, et al (2018) Genetic diversity of Deformed Wing Virus from Apis mellifera carnica (Hymenoptera: Apidae) and varroa mite (Mesostigmata: Varroidae). Journal of Economic Entomology https://doi.org/10.1093/jee/toy312
[16] Wu JY, et al (2011) Sub-lethal effects of pesticide residues in brood comb on worker honey bee (Apis mellifera) development and longevity. PLoS ONE 6(2): e14720.
[17] Di Prisco, G, et al (2013) Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc Natl Acad Sci 110(46): 18466–18471.
It’s “Common Knowledge” that Nosema Causes Dysentery…
But is it actually true?
Randy Oliver
ScientificBeekeeping.com
Yesterday I inspected a group of 40 hives returning from almond pollination. All were strong and healthy, other than two unusual deadouts, which no longer had any bees on the combs, and only a scattering of a few dead workers on the bottom boards. One hive had clear signs of dysentery on the top bars; the other didn’t. Upon further inspection, there were small arcs of apparently-healthy sealed brood remaining in both hives, and surprisingly, fresh eggs in the cells below that brood. What could be the cause of these unusual signs?
From my experience, the field signs above point the finger at nosemosis — as described by The World Organisation for Animal Health (OIE) [[1]]:
In a typical case of a colony being depleted because of a Nosema infection, the queen can be observed surrounded by a few bees, confusedly attending to brood that is already sealed.
Exactly as in the photo below, which I took during the CCD epidemic in the early 2000s.
Figure 1. Ten years ago, many beekeepers reported unexplained Colony Collapse Disorder, not associated with varroa. Colonies would quickly dwindle to a “silver dollar sized patch of bees and a queen,” as in this photo. Often times there would be patches of healthy brood remaining after the bees disappeared. I strongly suspect that a large part of the CCD epidemic was a result of the invasive wave of Nosema ceranae, as a part of a “perfect storm” that included evolving viruses, failure of varroacides, and evolving strains of European Foulbrood.
Practical application: Colonies that exhibit healthy brood during cool springtime weather, yet fail to build up, or in which the adult population appears to be declining, may well be suffering from nosemosis.
Nowadays, most of what is now termed “Colony Collapse” is caused by varroa and Deformed Wing Virus (DWV), and can be easily differentiated from collapse due to nosema by the telltale signs of brood dying from the virus, as well as the presence of mite fecal deposits on the ceilings of the cells (Fig. 2).
Figure 2. A typical varroa/DWV deadout, which typically occurs late in the season. Some cells contain dead pupae, or fully-developed adults that were unable to emerge. Note the distinctive traces left by varroa — the white fecal deposits at the top of the cells.
Practical application: Before you blame the collapse of your hive on something else, first pull a central brood frame out of the deadout, and check for the signs above.
But I still occasionally see colonies collapsing from nosema in early springtime. So how about my two deadouts — could nosema have been the cause? And why would only one hive exhibit dysentery? Again from OIE:
In some acute cases [of nosemosis], brown faecal marks are seen on the comb and the front of the hive, with sick or dead bees in the vicinity of the hive. However, the majority of colonies show no obvious signs of infection, even when the disease is sufficient to cause significant losses in honey production and pollination efficiency.
Practical application: So how could I tell whether nosema was the culprit? There’s only one way to tell whether a colony is infected by nosema ― I’ll put it in large type:
The only way for a beekeeper to diagnose nosema infection is by microscopy.
And sure enough, when I crushed dead bee samples from each of the bottom boards, Nosema ceranae spores were clearly evident and in abundance (Fig. 3).
Figure 3. Nosema spores ― the numerous small glowing elongated ovals — from a diluted sample of crushed bees viewed at 400x. The sample from the hive with dysentery looked identical to the one without.
Practical application: Get a microscope, or encourage your local club to purchase one. My favorite for viewing nosema is the Omano 36 [[2]], but even a cheap secondhand ‘scope will do.
So, yes to nosema, but no to having a correlation with dysentery. In fact, I’ve yet to find a single study that has ever demonstrated that nosema ― whether N. apis or N. ceranae — is responsible for causing dysentery.
A request and challenge: If you can find any study that has experimentally confirmed that nosema causes dysentery, please forward it to me.
What is dysentery?
We’ve all seen it ― dysentery on the front of a hive. Clearly those bees “really had to go” and let loose the moment they took off (Fig. 4).
Figure 4. The telltale signs of dysentery. Even worse is when it’s all over the top bars inside the hive, since it could then quickly spread any gut pathogens or toxic substances throughout the cluster. Photo credit: Monique Vescia
Honey bee workers are remarkably fastidious about hygiene, and do everything possible to avoid defecating within the hive. Foragers wait until they’re on the wing, whereas non-foraging “house bees” take “cleansing flights” when necessary.
Many of us have had to apologize to our neighbors about the orange polka dots on their cars or clotheslines. Although those drops of bee poo are normally a mere annoyance, back during the Viet Nam War, it became an international issue, when accusations were leveled about purported chemical warfare — the evidence being spots of “yellow rain” dotting leaves [[3]]. Such “rain” would have been immediately recognized by any beekeeper who’s enjoyed the experience of walking in an apiary when the sun comes out after the bees had been confined by weather to their hives for several days.
During periods of forced confinement, honey bees have a remarkable ability to “hold it” in order to avoid soiling their nest (for months if necessary). A bee’s rectum has the ability to distend to the point that it nearly fills the abdomen (Fig. 5).
Practical application: Dysentery is an indication that a colony has a serious problem ― one that sometimes leads to the death of the hive, but not necessarily from nosema. I will later return to the subject of the causes of dysentery.
So why does everyone think that nosema causes dysentery?
Here’s where we get into one of my pet peeves about honey bee popular books, as well as scientific papers ― people tend to repeat something they’ve heard just because it seems to make sense — but without first actually checking the facts.
I see it in scientific paper after paper. The author says that dysentery is a “sign of nosema” and throws in a citation to support that claim. I’ve checked dozens of those citations, and have yet to find a single one that actually refers to any study that actually demonstrated that nosema infection induced dysentery. Keep in mind that nosema infects the cells that line the bee’s midgut; the “environmental spores” that are found in the hindgut have never been shown to infect or irritate the lining of the rectum. And if nosema did indeed cause lack of hygienic control, you’d expect to see the fecal contents leaking out as little dribbles, not in the huge streaks from a rectum able to be kept full to bursting.
Unfortunately, many authors don’t bother to check, and something that someone once said gets repeated over and over again until it becomes “common knowledge.”
Practical application: This is not the only “common knowledge” myth in the bee literature that possibly needs to be debunked ― I’ll be addressing others in the future.
The sad part
The really sad thing about this misconception is that researchers and textbook authors have no excuse for repeating such an assumption, since the lack of nosema-induced dysentery was well established by 1922, and the cause of dysentery by 1935. But I’ve rarely seen any of the many recent papers on nosema cite those nicely-aged, but scientifically accurate studies.
Practical application: Not every author or researcher does their homework. I’ve learned over the years to double check the supportive citations in scientific papers myself, a process that often leads me down a string of misinterpretations (and sometimes to different conclusions than reached by the authors) [[5]]. I’m often appalled by the amount of scientific sloppiness allowed to get by the peer reviewers of papers.
Nosema apparently does not cause dysentery
I discovered this when I began deep research into N. ceranae in 2006, and found that most everything that we needed to know about nosema had been clearly explained in a forgotten USDA bulletin published in 1919 [[6]]. I love to read these old studies, and am often impressed by the scientific diligence of the government-funded authors — in this case a G.F. White spent 9 years experimenting with Nosema apis in every way. White’s 58-page summary, in my opinion, still stands as perhaps the most informative study on the parasite, and the disease it may cause, yet published in the English language.
Practical application: One thing that I found was that N. ceranae was not all that different from N. apis. Once the invasive wave of N. ceranae passed through North America in the 2000s, the main differences that I now see between the two species is that ceranae may crop up under certain conditions during warm weather, its spores are far less cold tolerant, and its spore counts may be much higher (although they do not cause a whitening of the hindgut, as with N. apis). Both species are common parasites of the midgut in springtime (with ceranae now being far more prevalent than apis), generally appear to be rather harmless; but may cause disease in stressed colonies.
Allow me to quote Dr. White:
“Care should be observed that Nosema-disease is not confused with dysentery…In fact the two disorders are very different and should be considered, for the present at least, as having no direct relation to each other. As both conditions are widely distributed and occur most frequently in the spring of the year. It is to be expected that not infrequently both of them may be encountered together in the same colony.”
The start of the “association” between nosema and dysentery is perhaps explained by famed bee pathologist Dr. Leslie Bailey [[7]]:
Infection by Nosema apis is commonly believed to make honeybees (Apis mellifera) void faeces in and on the colony instead of away in flight — a condition known as ‘dysentery’ by beekeepers. Evidence that seemed to support this was provided by Lotmar (1951), who found more faeces accumulated in caged infected bees than in healthy ones. No field experiments have shown this effect…
Dysentery was not caused primarily by N. apis, because it occurred whether colonies were severely infected or not…Most of the colonies that survived managed to clear the faeces away; the few still with combs contaminated with faeces were among those with more than 25% of bees infected… If infection is spread by dysentery, this is not unexpected, since infection would spread most in colonies that suffered most dysentery; but if infection causes dysentery a more striking correlation between dysentery and severe infection would be expected than actually occurred.
Practical application: Let me make clear that should a bee already infected with nosema happen to defecate within the hive due to some other gut issue, it will certainly transmit nosema spores to the other bees that clean up the mess. But the dysentery was apparently the result of something other than nosema itself.
Part of the apparent confusion between nosema and dysentery may be due to the fact that they both commonly occur in early spring. In my own apiaries, dysentery comes and goes in the springtime, but after checking numerous dysentery samples (from my own apiaries and numerous samples from others) under the microscope, I have yet to notice any correlation between nosema and dysentery.
Biological evaluation: One could reasonably expect a gut parasite to induce dysentery in its host, as a means of transmission of its spores. But if nosema did indeed induce dysentery, every infected colony would quickly be overwhelmed by spore transmission, and die before spring. This we do not see. Could it be that both species of nosema are actually rather benign parasites, which under “normal” circumstances do not cause appreciable harm to the colony? That is, unless the colony suffers from dysentery due to some other reason.
In France, the impact upon colonies from N. ceranae has been referred to as “dry nosemosis,” since it is clear that the parasite doesn’t cause dysentery. Perhaps we should now question whether N. apis ever did either.
Next
In my next two articles I’ll discuss our current state of knowledge about Nosema ceranae ― its seasonality (and the causes for that seasonality), its effects upon colony performance (or lack thereof), and the best ways to monitor for it.
And then I’ll follow with an article about the potential causes of dysentery in bees.
Beekeeping is more difficult today than it used to be.
Our changing agricultural landscape provides less forage, and growers still apply pesticides to freely (although the pesticide situation for bees today is far better than it used to be in the ’60s and ’70s).
The main problem for honey bees worldwide is the recent invasion of the varroa mite, which acts as a vector for Deformed Wing Virus.
The long-term solution is to breed bees naturally resistant to the mite. In this presentation I offer a brief version of how to go about doing it. There is more information at my website.
Until such bee stock is more widely available, good bee husbandry requires occasional treatments to control the mite. My sons and I run a successful commercial beekeeping operation, and have used only organically-approved treatments since 2001.
I also give a progress report on our registration of extended-release oxalic acid for mite control. This organic treatment will help us to keep healthy, thriving bees.
I created a handy spreadsheet in Excel for calculating Henderson-Tilton efficacies. Here’s a snip of it:
If you wish, you can enter your raw data into the blue unprotected grid, and then calculate the means or medians (and standard errors of the means if you wish to display them) there. Then enter those calculated values into the yellow cells (erase the existing illustrative numbers). The easiest way is to enter into each yellow cell “=O27” if, for example, the mean value is in cell O27.
Or, if you’ve already calculated those values, simply enter them into the yellow cells.
Everything on this website is open access and freely given to beekeepers and researchers worldwide, on a not-for-profit basis. I gladly accept gifts of appreciation from my readers and supporters. You can donate via Paypal below. Notice: I will, for tax purposes, treat all Paypal donations as "gifted research grants" to be applied towards beekeeping research and the reporting on thereof -- given by you with "detached and disinterested generosity" out of "affection, respect, admiration, charity or like impulses." Since I am not a 501(c)(3), your gifts are not tax deductible.
Thank you!
Randy
Supporters
ScientificBeekeeping is a not-for-profit enterprise, and I'm happy to receive notes of thanks for how information on this site has contributed to my readers' success at beekeeping (and sometimes saved them hundreds or thousands of dollars). It is your support that allows me to devote my life to this site. All donations go towards website maintenance, bee research costs (typically tens of thousands of dollars per year), re-donations to fund research by others, and a small amount to partially offset the huge number of hours that I spend in research and writing. I guarantee that every penny is pinched and well spent!
I appreciate the notes of support that I receive from all over the world. Seeing who is donating, their locations, and the little notes that come with contributions help to let me know how I should direct my research and writing. In appreciation, and in order to show who is supporting my research and writing, I honor below those who have recently made substantial donations:
Recent $1000-plus MAJOR SUPPORTERS
Dylan Kelly AgPollen.com
BuzzsBees, California
South Jersey Branch of the NJBA
Mason Valley Beekeepers, Nevada
Northeast Oklahoma Beekeepers Assoc.
Ramsey Wilson
Lauri Sandquist, Seattle
Hiatt Honey, Washington
Miller Honey Mandarins, California
Paul Hosticka, Washington
William Boytim, Texas
Mt. Diablo Beekeepers Association, California
Chester County Beekeepers Association, Pennsylvania
Tualatin Valley Beekeepers Association, Oregon
Beekeepers Association of the Ozarks
Ron Davis, Virginia
Michigan Commercial Beekeepers Association
Help Save the Bees Foundation, Nevada
Ludwig Armbruster Imkerschule, Weimar, Germany
Northeast Oklahoma Beekeepers Associatioin
South Jersey Branch of New Jersey Beekeepers Association








 Figure 5. Move fast, but don’t rush! My sons found an exceptionally productive hive last season, and suggested that I check it for mites. It was late on a cold day, so I hurriedly took a sample. To my horror, as soon as I dumped the bees into the alcohol, my eye spotted the queen’s abdomen. Unfortunately, there’s no way to save a queen once she gets immersed. The mite wash turned out to be zero, so I immediately grafted 100 daughter larvae in order to save her genetics.
Figure 5. Move fast, but don’t rush! My sons found an exceptionally productive hive last season, and suggested that I check it for mites. It was late on a cold day, so I hurriedly took a sample. To my horror, as soon as I dumped the bees into the alcohol, my eye spotted the queen’s abdomen. Unfortunately, there’s no way to save a queen once she gets immersed. The mite wash turned out to be zero, so I immediately grafted 100 daughter larvae in order to save her genetics.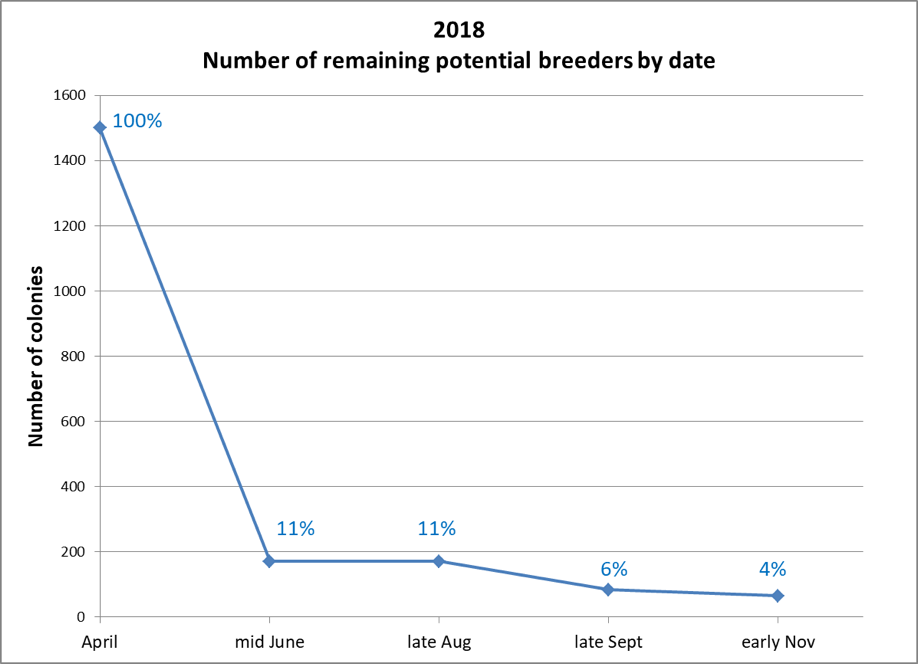


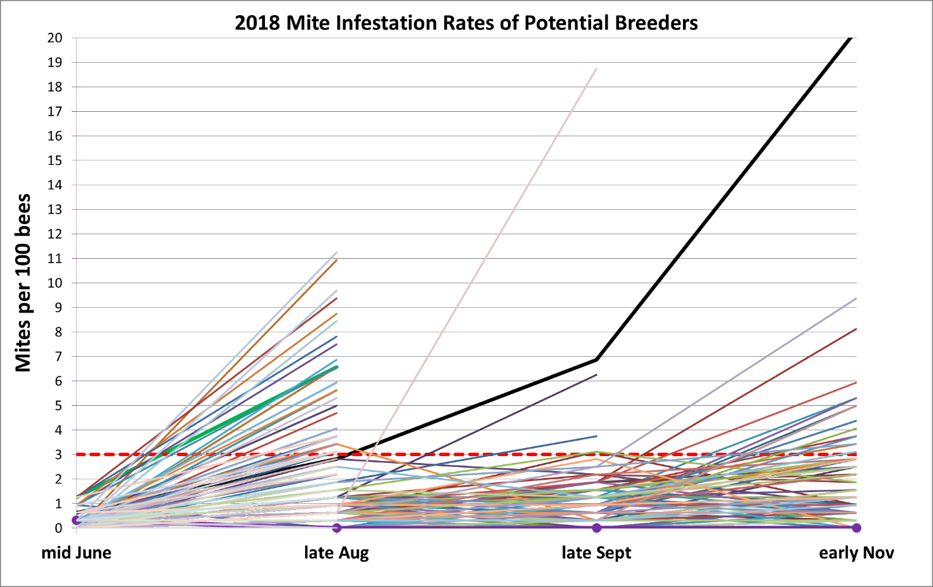


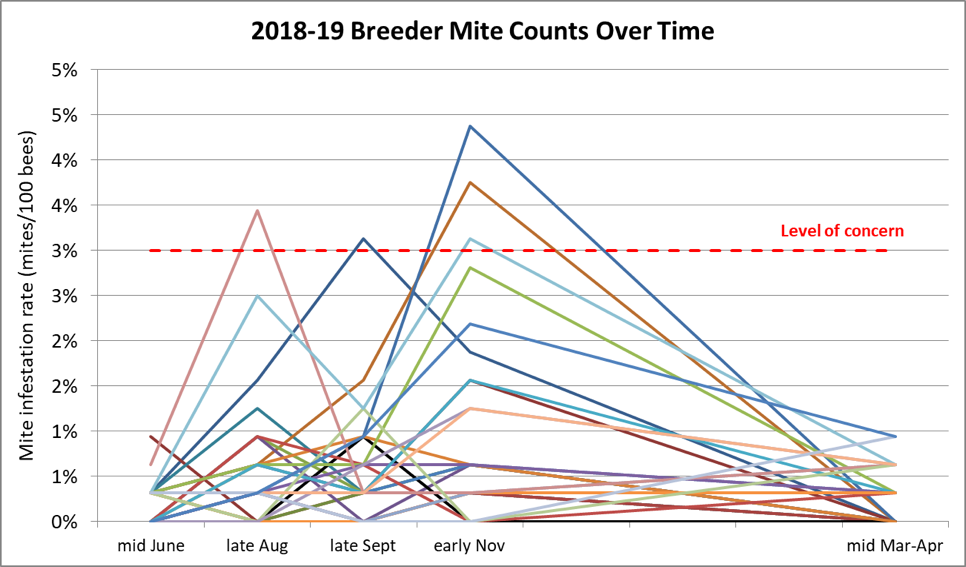
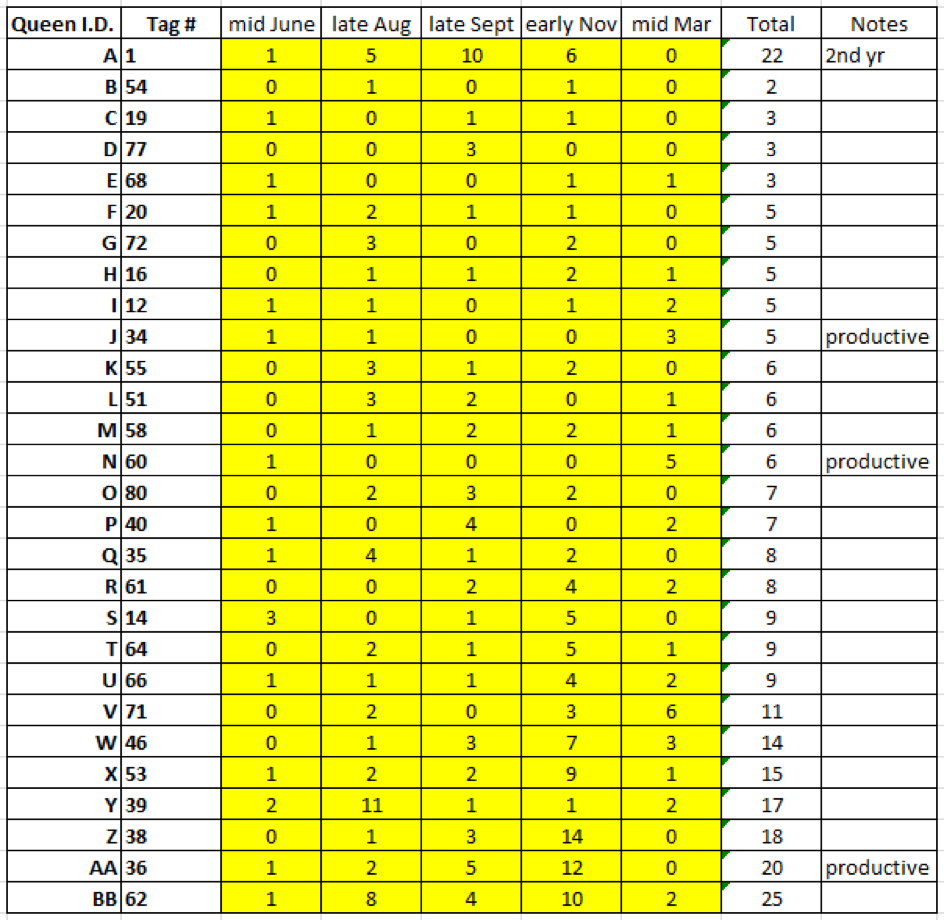
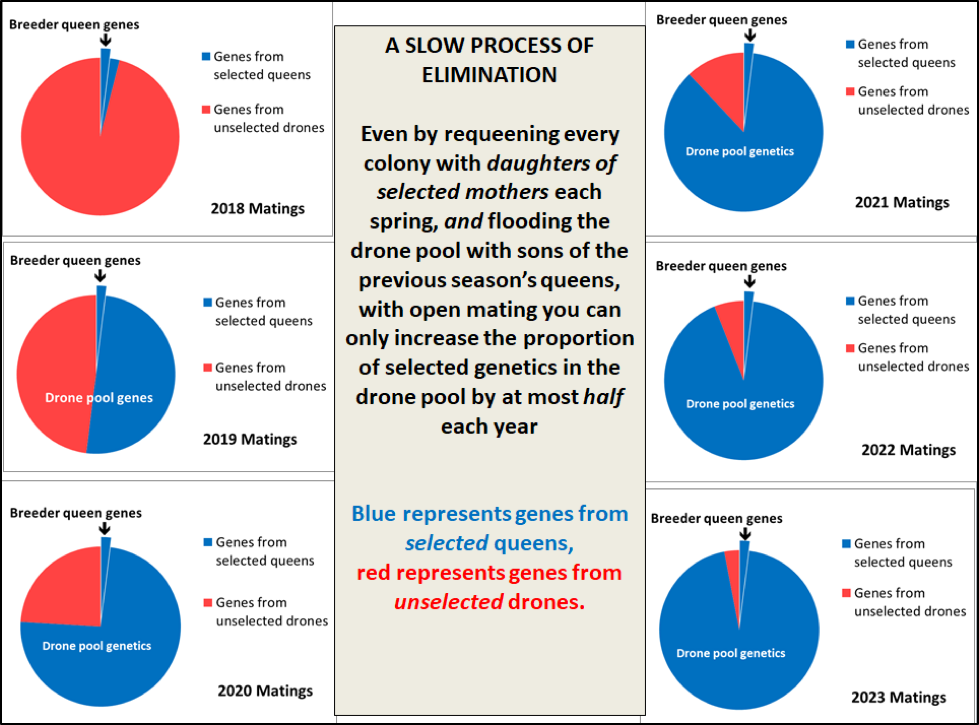


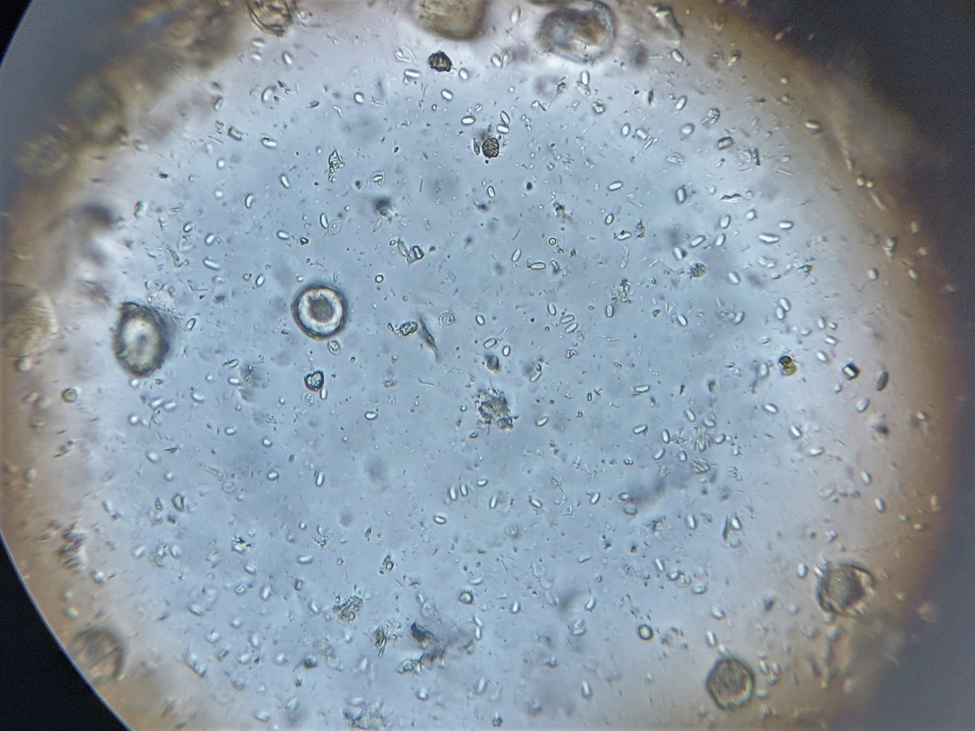

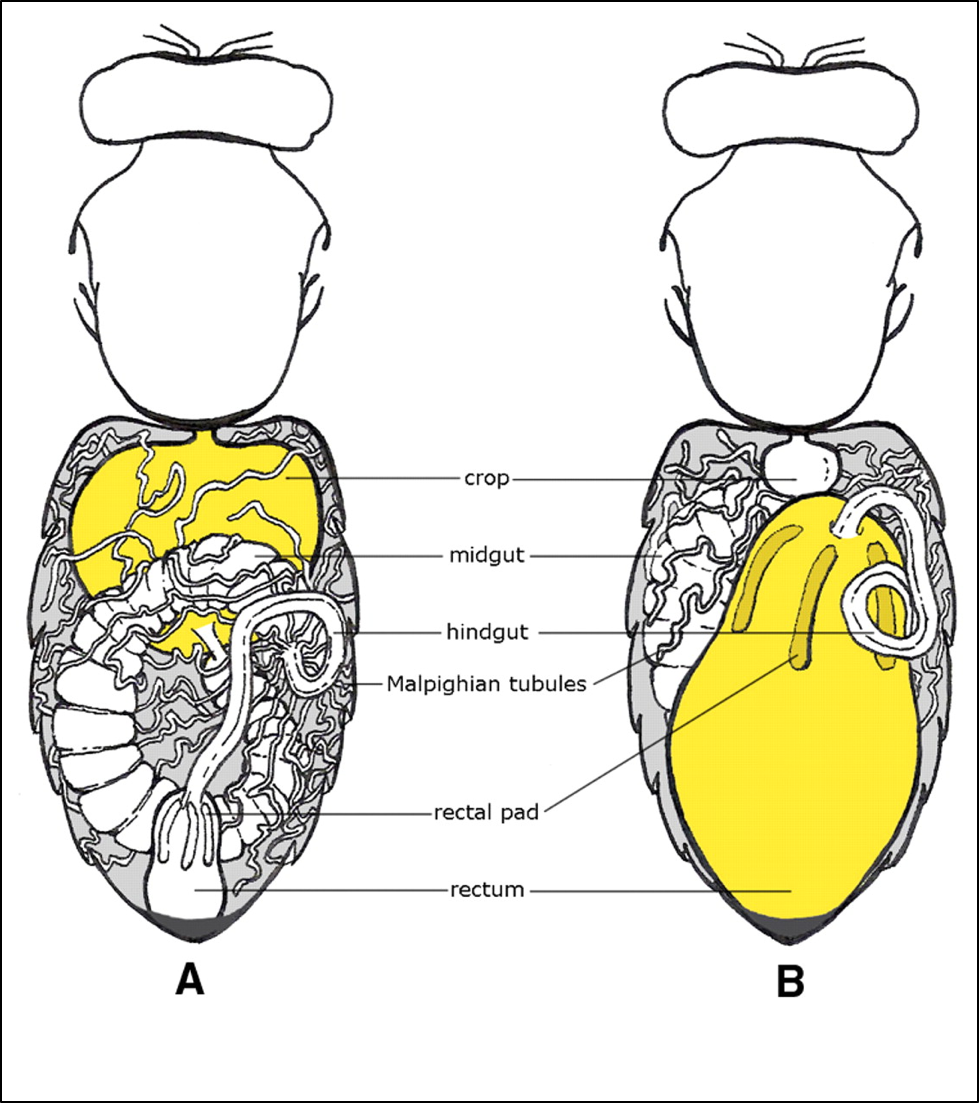


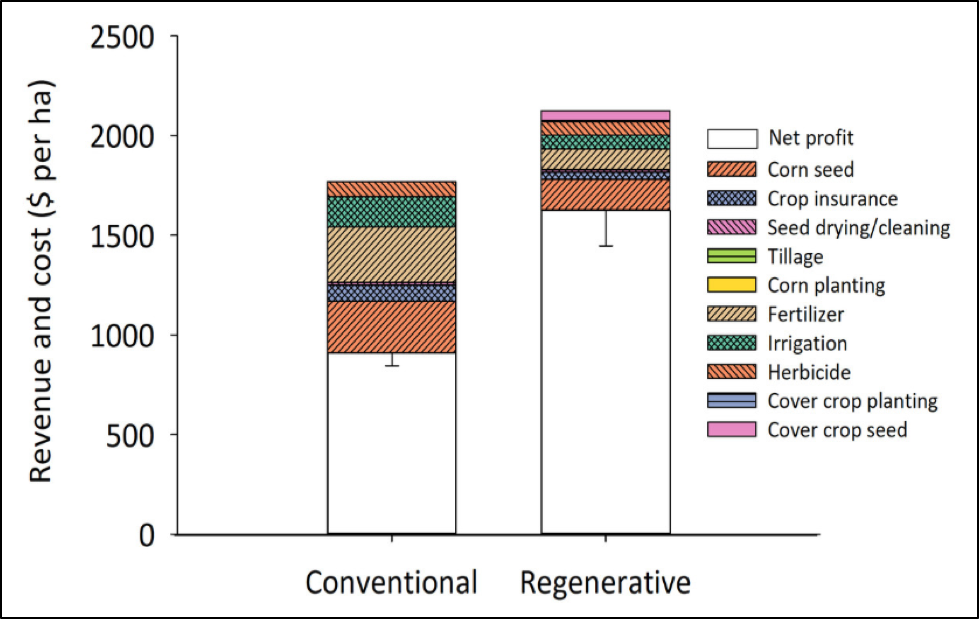


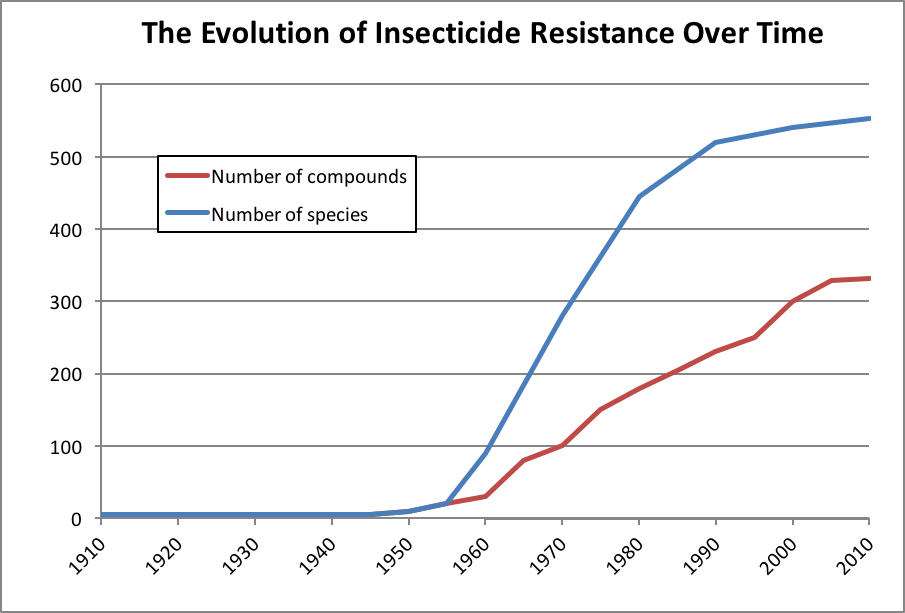
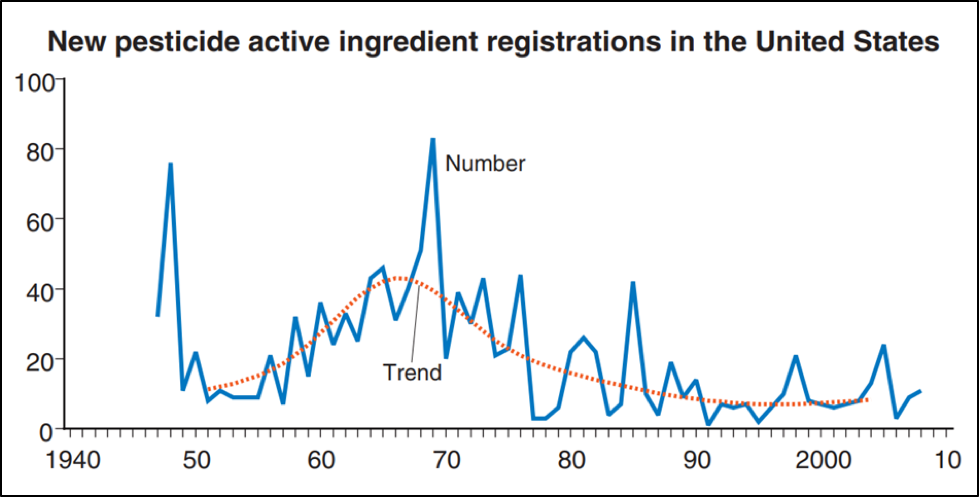
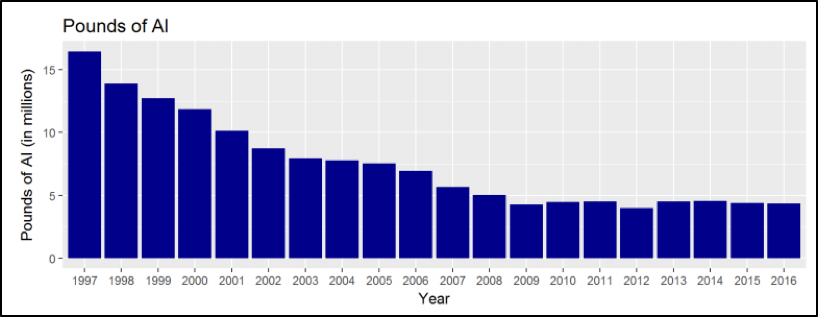
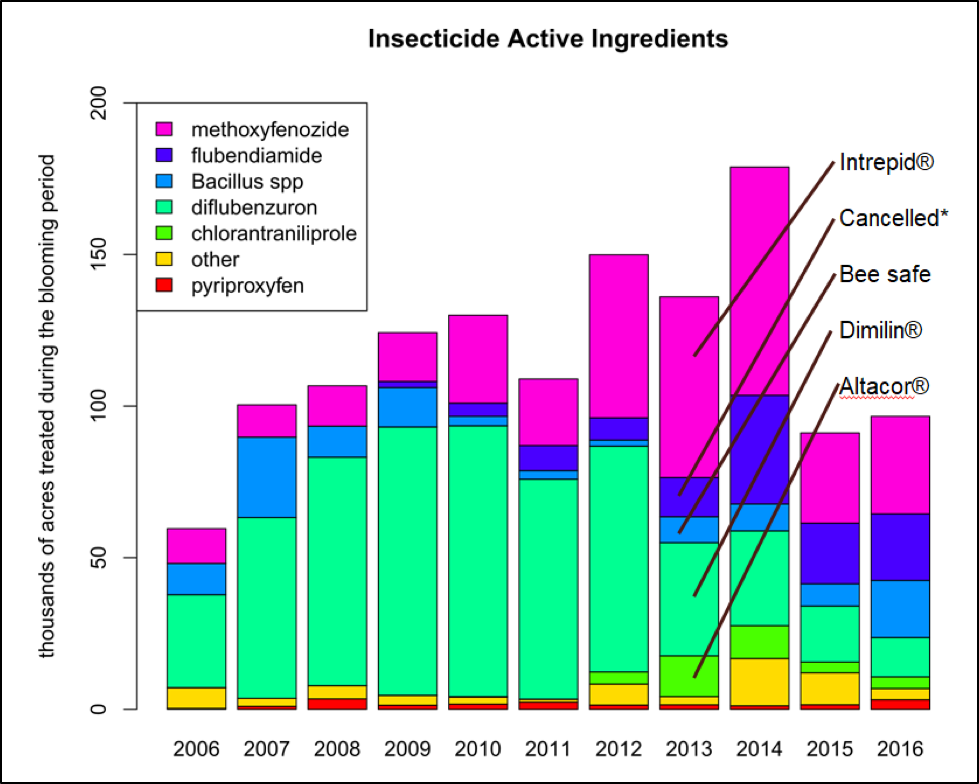


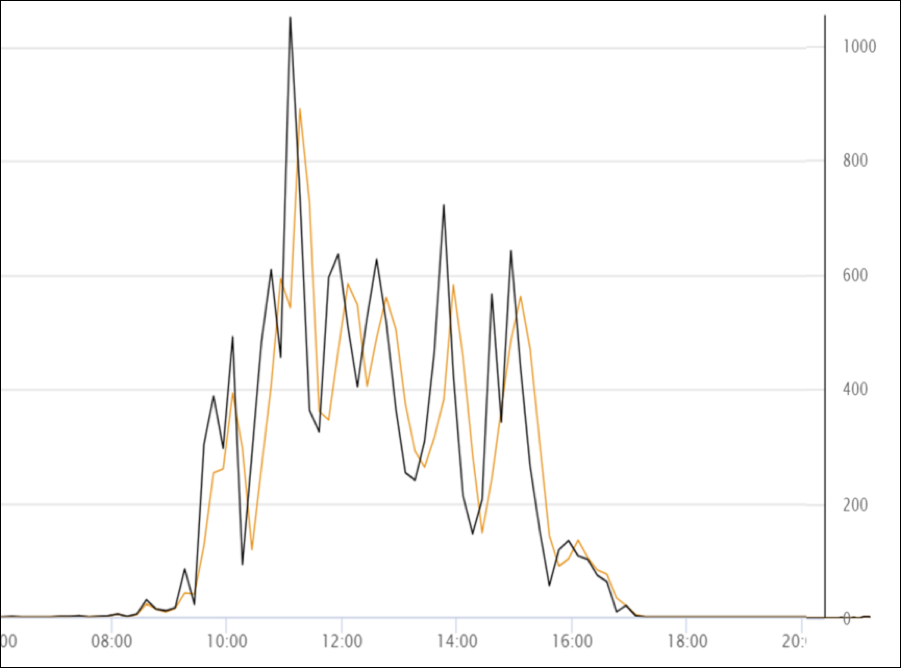



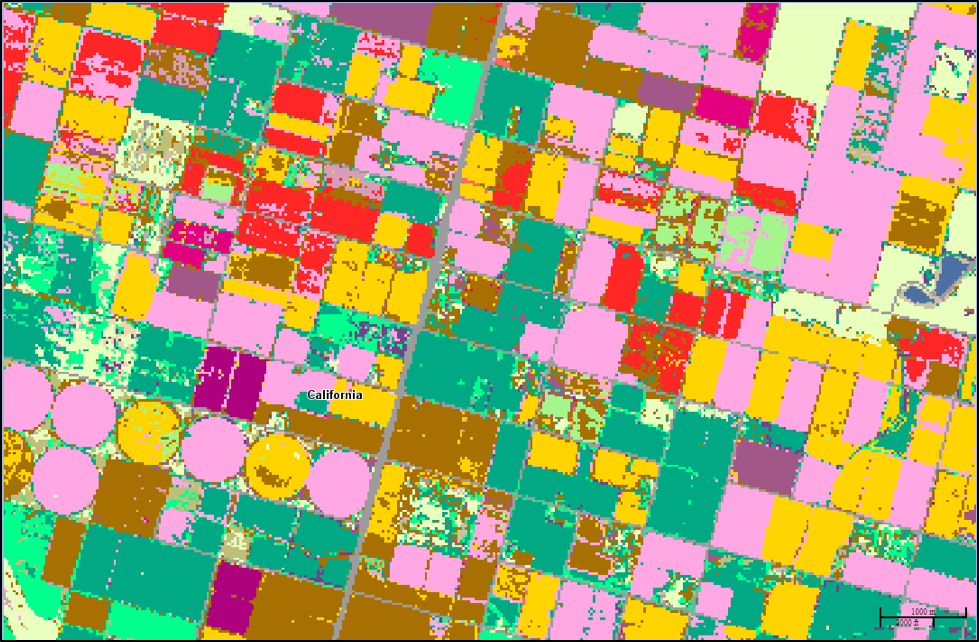


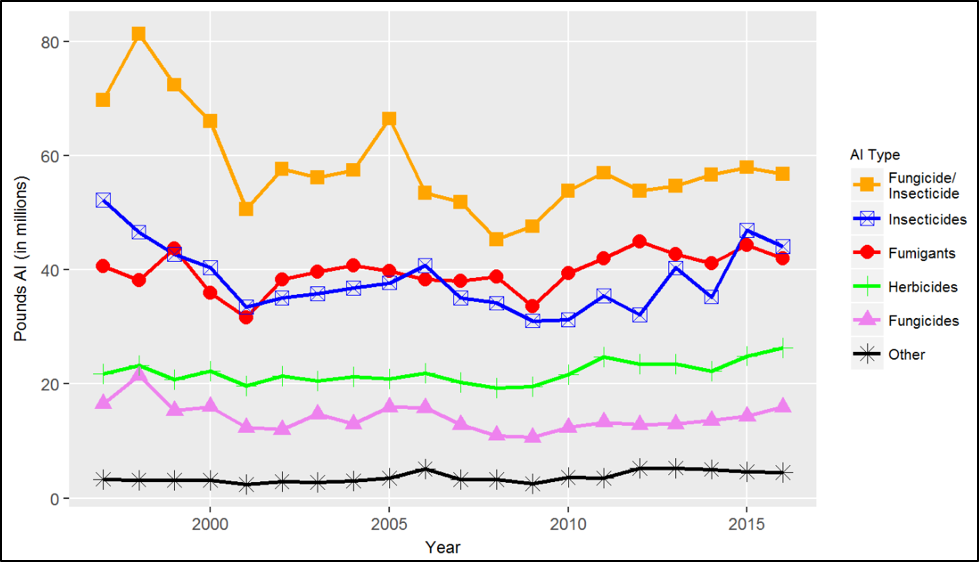
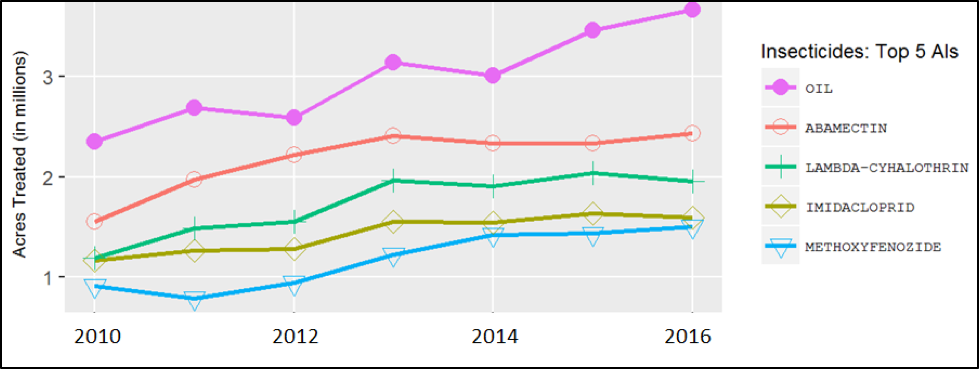
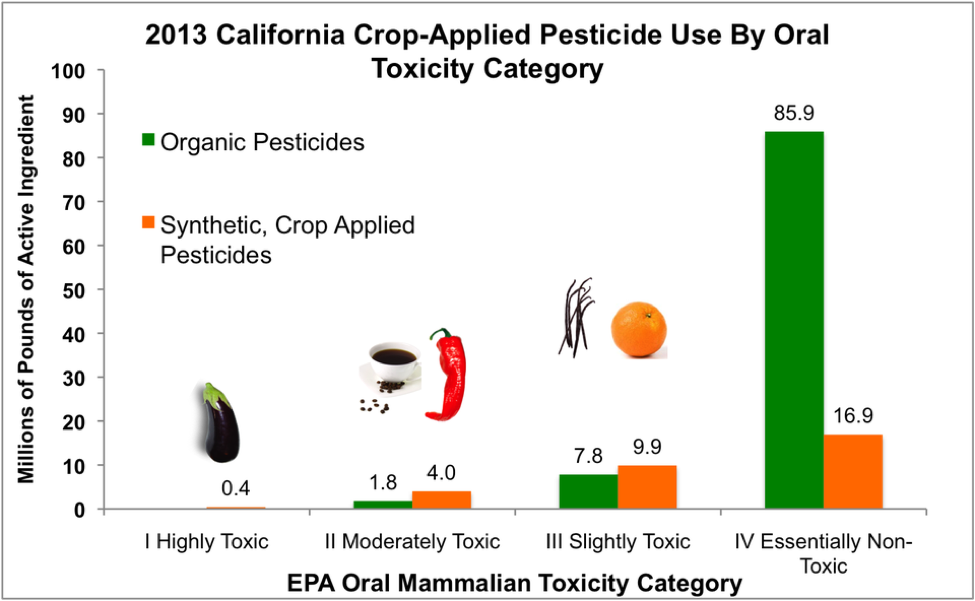


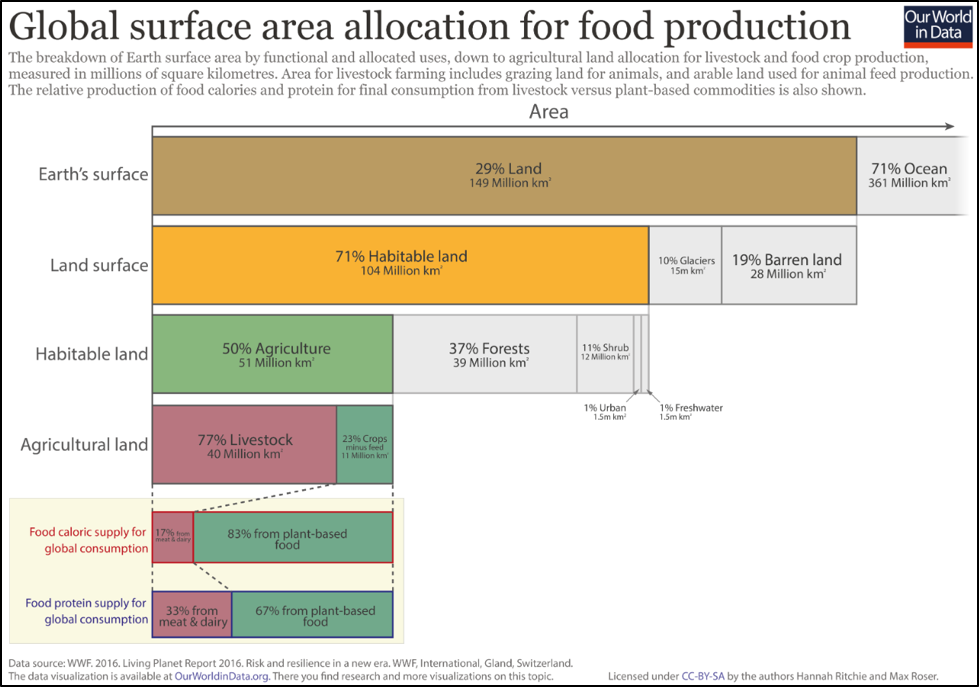
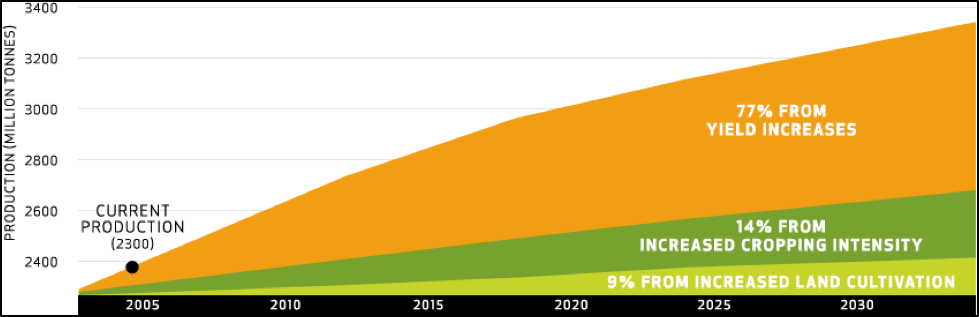



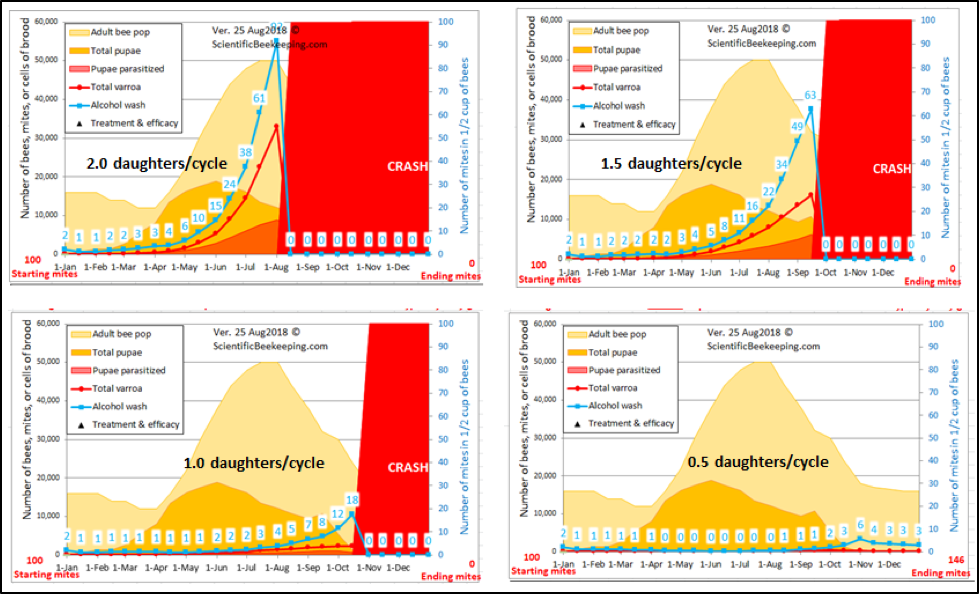








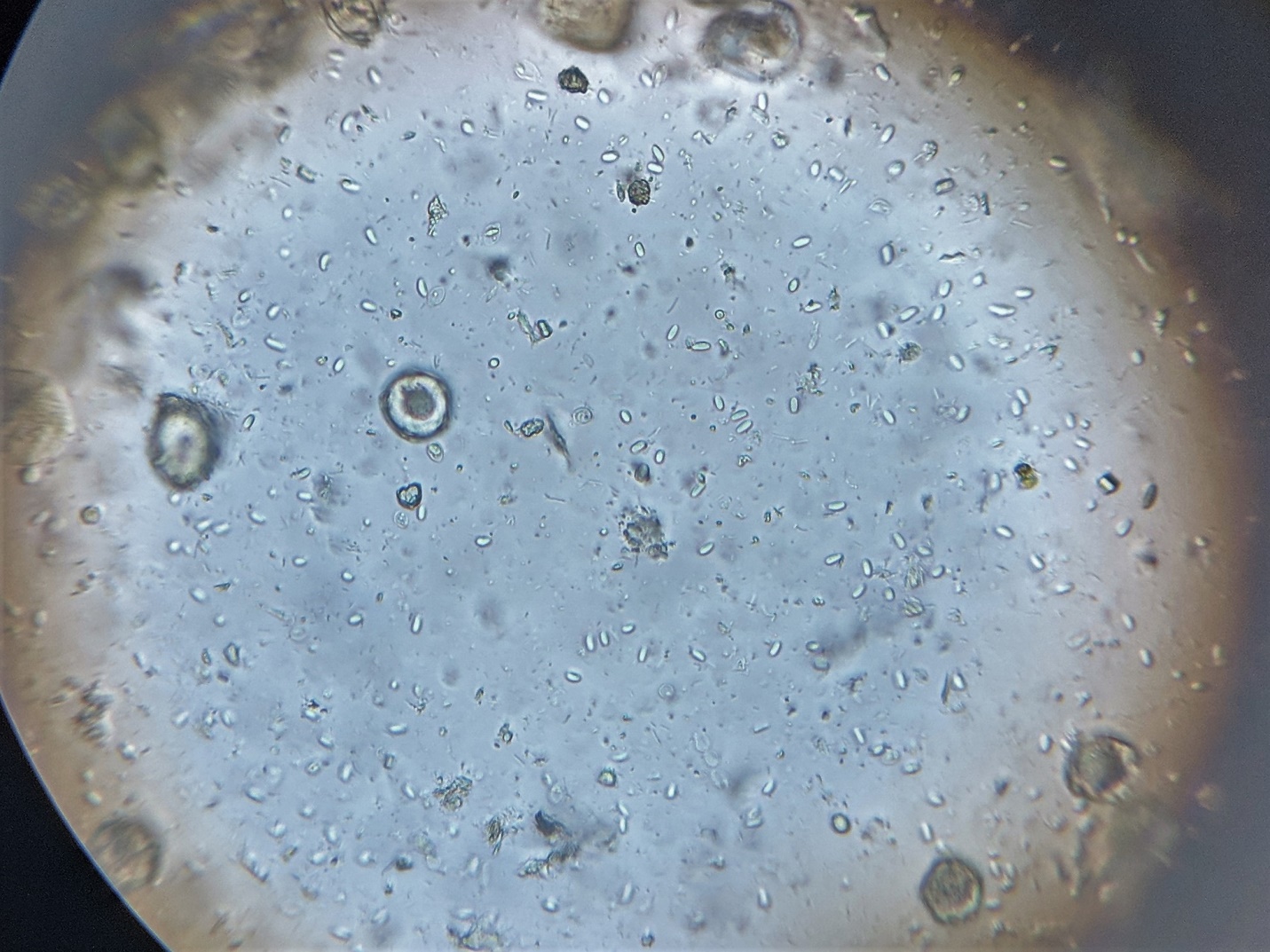

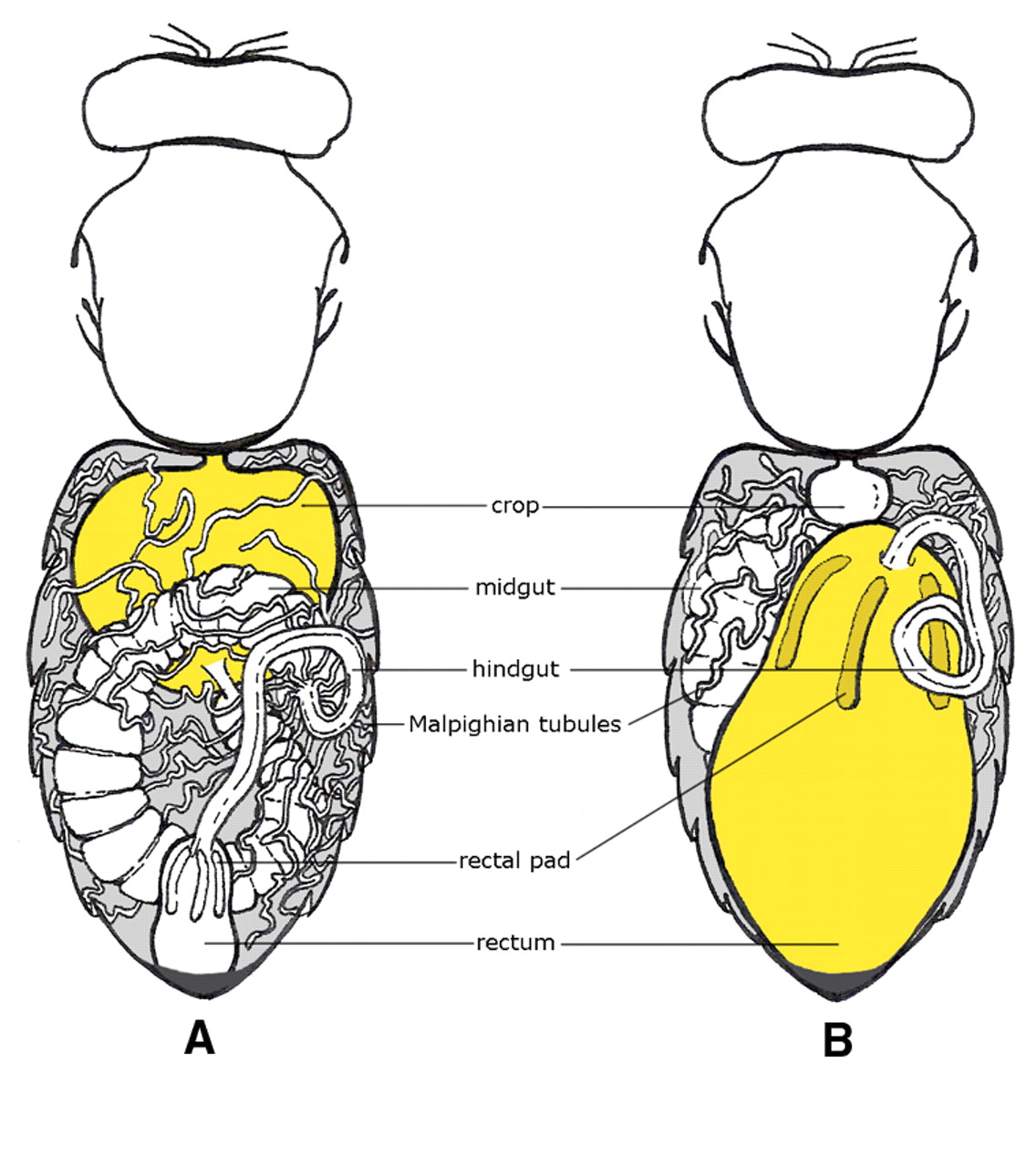

{kind=link}